
Презентация по математике НОК (6 класс) доклад, проект
Наименьшее общее кратное
Туда и обратно
идет 3 ч.
Туда и обратно
идет 2 ч.
От одной пристани к другой ходят два катера. Начинают работу одновременно в 8 ч утра. Первый катер на рейс туда и обратно тратит 2ч, а второй — 3 ч.
Через какое наименьшее время оба катера опять окажутся на первой пристани, и сколько рейсов за это время сделает каждый катер?
Сколько раз за сутки эти катера встретятся на первой пристани, и в какое время это будет происходить?
Искомое время должно делиться без остатка и на 2, и на 3 то есть должно быть кратным числам 2 и 3.
Числа кратные:
2:
2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24.
3:
3, 6, 9, 12, 15, 18, 21, 24.
Подчеркнем общие кратные чисел 2 и 3.
Наименьшее общее кратное
Значит, через 6 ч после начала работы два катера одновременно окажутся на первой пристани.
Сколько рейсов за это время сделает каждый катер?
Первый – 3 рейса, второй – 2 рейса.
Сколько раз за сутки эти катера встретятся?
4 раза
В какое время это будет происходить?
В 14 ч, 20 ч, в 2 ч ночи, в 8 утра.
Обозначение: НОК (2; 3) = 6.
Алгоритм нахождения НОК:
Пример:
Найдите наименьшее общее кратное чисел: 75 и 60.
75 = 3 ∙ 5∙ 5
60 = 2∙ 2∙ 3 ∙ 5
НОК (75; 60) = 3 ∙ 5 ∙ 5∙ 2∙ 2 = 75∙ 2 ∙ 2 = 300.
1. Разложить все числа на простые множители.
2. Написать разложение одного из чисел (лучше наибольшего).
3. Дополнить данное разложение теми множителями из разложения других чисел, которые не вошли в написанное разложение.
Определение
Наименьшее натуральное число, которое делится на каждое из данных натуральных чисел, называется наименьшим общим кратным.
Нужно найти НОК чисел 45 и 60.
Вдоль дороги от пункта А поставлены столбы через каждые 45 м. Эти столбы решили заменить другими, поставив их на расстоянии 60 м друг от друга. Найдите расстояние от пункта А до ближайшего столба, который будет стоять на месте старого.
45 = 2 ∙ 3 ∙ 5 60 = 2 ∙ 5 ∙ 2 ∙ 2
НОК (45; 60) = 60 ∙ 3 = 180,
значит 180 м.
Ответ: 180 м.
15 = 3 ∙ 5; 20 = 2 ∙ 2 ∙ 5;
12 = 2 ∙ 2 ∙ 3;
НОК (15; 20; 12) = 20 ∙ 3 = 60, следовательно, через 60 суток.
В портовом городе начинаются три туристских теплоходных рейса, первый из которых длится 15 суток, второй 20 суток и третий 12 суток. Вернувшись в порт, теплоходы в этот же день снова отправляются в рейс. Сегодня из порта вышли теплоходы по всем трем маршрутам. Через сколько суток они впервые снова вместе уйдут в плавание?
Сегодня из порта вышли теплоходы по всем трем маршрутам. Через сколько суток они впервые снова вместе уйдут в плавание?
Нужно найти НОК чисел 15; 20 и 12.
Слово «крат» — старинное русское слово (XI век), означающее «раз». Слова «многократно» означает «много раз».
Понятием кратного пользуются в жизненной практике при установлении вида года. Через каждые три обыкновенных года, в каждом из которых по 365 дней (в феврале 28 дней), бывает четвертый год, так называемый високосный, в котором 366 дней (в феврале 29 дней).
Если число, которым выражается указанный год, есть число, кратное 4, то указанный год високосный, а если не кратно 4, то год обыкновенный. Так, 2016 год — високосный, так как 2016 кратно 4, 2017 — не високосный, так как 2017 не кратно 4.
Историческая минутка
а) НОК (а; b) = 2 ∙ 2 ∙ 5 ∙ 5 ∙ 11 ∙ 3 = 3300;
НОК (а; b) = 2 ∙ 2 ∙ 5 ∙ 5 ∙ 7 = 700
б)Так как b делится на а, то НОК будет само число b;
б) НОК (12; 16) = 2 ∙ 2 ∙ 2 ∙ 2 ∙ 3 = 48;
Найдите наименьшее общее кратное чисел:
а)12 и 16;
I
12 = 2 ∙ 2 ∙ 3; 16 = 2 ∙ 2 ∙ 2 ∙ 2
Найдите наименьшее общее кратное чисел:
б) 396 и 180;
г) НОК (396; 180) = 2 ∙ 2 ∙ 3 ∙ 3 ∙ 11 ∙ 5 = 1980;
II
396 = 2 ∙ 2 ∙ 3 ∙ 3 · 11; 180 = 2 ∙ 2 ∙ 3 ∙ 3 · 5
в) 168, 231 и 60.
Найдите наименьшее общее кратное чисел:
е) НОК (168; 231; 60) = 231 ∙ 2 ∙ 2 ∙ 2 ∙ 3 · 7 · 5 = 194 040
III
168 = 2 ∙ 2 ∙ 2 ∙ 3 · 7; 231 = 1 · 231; 60 = 2 ∙ 2 ∙ 3 ∙ 5
Домашнее задание:
п. 7
№ 202 (б, г)
№ 203
№ 204
48 см
40 см
?
?
НОД чисел 48 и 40 — длина стороны квадрата
№ 191. Лист картона имеет форму прямоугольника, длина которого 48 см, а ширина 40 см. Этот лист надо разрезать без отходов на равные квадраты. Какие наибольшие квадраты можно получить из этого листа?
НОД (48; 40) = 8,
значит, квадраты будут со стороной 8 см.
48 ∙ 40 = 1920 (см2) – площадь прямоугольника.
8 ∙ 8 = 64 (см²) — площадь квадрата,
1920 : 64 = 30 (квадратов)
Ответ: квадраты со стороной 8 см; 30 квадратов.
№ 182. Являются ли числа 54 и 65 взаимно простыми? Найдите наименьшее общее кратное чисел 54 и 65. Равно ли оно произведению 54 и 65? Запишите какие-нибудь два взаимно простых числа. Найдите наименьшее общее кратное этих чисел. Сделайте вывод.
Являются ли числа 54 и 65 взаимно простыми? Найдите наименьшее общее кратное чисел 54 и 65. Равно ли оно произведению 54 и 65? Запишите какие-нибудь два взаимно простых числа. Найдите наименьшее общее кратное этих чисел. Сделайте вывод.
54 = 2 ∙ 3 ∙ 3 ∙ 3
65 = 5 ∙ 13
НОК (54; 65) = 54 · 65 = 3510;
24 = 2 ∙ 2 ∙ 2 ∙ 3
35 = 5 · 7
НОК (24; 35) = 24 · 35 = 840;
Вывод:
Наименьшее общее кратное двух взаимно простых
чисел равно их произведению.
Найдем наименьшее общее кратное любых двух взаимно простых чисел
№ 183. Найдите наименьшее общее кратное чисел:
а) 45 и 135; б) 34 и 170.
Равно ли оно одному из данных чисел?
Так как большее число делится на меньшее, то наименьшим
общим кратным этих чисел будет являться большее число.
НОК (45; 135) = 135; НОК (34; 170) = 170.
Наибольший общий делитель этих чисел:
НОД (45; 135) = 45; НОД (34; 170) = 34.
НОД (22; 66) = 2 · 11 = 22
Найдите наименьшее общее кратное и наибольший общий делитель чисел 22 и 66:
НОК (22; 66) = ? НОД (22; 66) = ?
НОК (22; 66) = 2 · 3 · 11 = 66
I
НОД (39; 65) = 13.
Найдите наименьшее общее кратное и наибольший общий делитель чисел 39 и 65:
НОК (39; 65) = ? НОД (39; 65) = ?
НОК (39; 65) = 5 · 13 · 3 = 195;
II
В классе:
№ 181 (в, д, е)
№ 206 (а, в)
№ 207
Домашнее задание:
№ 202 (а, в)
№ 206 (а, в)
№ 207
Ответить на вопросы:
Какое число называют наименьшим общим кратным натуральных чисел а и b?
Какое число называют наибольшим общим делителем натуральных чисел а и b?
Какое число является наименьшим общим кратным чисел т и п, если число т кратно числу n?
Какие натуральные числа называют простыми?
Какие натуральные числа называют взаимно простыми?
Наименьшее общее кратное и наибольший общий делитель » задачи
НОД и НОК »
Наименьшее общее кратное(135,5)
Решение: Наиме́ньшее о́бщее кра́тное (НОК) двух целых чисел m и n есть наименьшее натуральное число, которое делится на m и n. Обозначается одним из следующих способов: исходя из этого наименьшое общее кратное чисел 135 и 5 это 135
Обозначается одним из следующих способов: исходя из этого наименьшое общее кратное чисел 135 и 5 это 135НОК (135,5)=3в кубе *5=27*5=135
Наименьшее общее кратное 35и 70
Решение: Чтобы найти НОК (a; b), нужно разложить данные числа на простые множители и найти произведение всех простых множителей, взятых с наибольшим показателем степени.
35 : 5 = 7 70 : 2 = 35
7 : 7 = 1 35 : 5 = 7
35 = 5 * 7 7 : 7 = 1
70 = 2 * 5 * 7
НОК (35 и 70) = 2 * 5 * 7 = 70 — наименьшее общее кратноеНайдите наименьшее обшее кратное 8и10 12и48 25и12 75и45
Решение: А) 40
б) 48
в) 300
г) 225Чтобы найти наименьшее общее кратное ( НОК) надо:
Чтобы найти наименьшее общее кратное нескольких натуральных чисел, надо:
1) разложить их на простые множители;
2) выписать множители, входящие в разложение одного из чисел;
3) добавить к ним недостающие множители из разложений
остальных чисел;
4) найти произведение получившихся множителей.
Применяя это правило решим примеры:
8 и 10
раскладываем 8- это 4 2 2
раскладываем 10 — это 5 2
теперь перемножаем : 4*2*2*2*5 = 160
НОК 8 и 10 = 160
И так далее:
12 и 48
12- 6 2
48 — 8 2 3
6*2*8*2*3= 576 НОК 12 и 48 = 576
25 и 12
25-5 5
12 — 6 2
5*5*6*2= 300 НОК 25 и 12= 300
75 и 45
75- 5 5 3
45 — 5 3 3
5*5*3*5*3*3= 3375 НОК 75 и 45 = 3375Найди наименьшее общее кратное (НОК)
1) 12 и15
2) 20 и 35
3) 16 и 20
4) 15 и 18
Решение: Чтобы найти НОК нескольких чисел, нужно разложить эти числа на простые множители и найти произведение всех простых множителей, взятых с наибольшим показателем степени.
1) НОК (12 и 15) = 2 * 2 * 3 * 5 = 60 — наименьшее общее кратное
12 = 2 * 2 * 3 60 : 12 = 5
15 = 3 * 5 60 : 15 = 4
2) НОК (20 и 35) = 2 * 2 * 5 * 7 = 140 — наименьшее общее кратное
20 = 2 * 2 * 5 140 : 20 = 7
35 = 5 * 7 140 : 35 = 4
3) НОК (16 и 20) = 2 * 2 * 2 * 2 * 5 = 80 — наименьшее общее кратное
16 = 2 * 2 * 2 * 2 80 : 16 = 5
20 = 2 * 2 * 5 80 : 20 = 4
4) НОК (15 и 18) = 2 * 3 * 3 * 5 = 90 — наименьшее общее кратное
15 = 3 * 5 90 : 15 = 6
18 = 2 * 3 * 3 90 : 18 = 5Найди.
( НОК- наименьшее общее кратное) А) НОК(162,216). Б) НОК (350,400) В) НОК(60,220). Г) НОК(23,47) Д) НОК(495,33). Е) НОК(300,270) Ж) НОК(441, 4410). З) НОК(68,102,136)
Решение: А) НОК(162, 216)=2*2*2*3*3*3*3=648
162= 2 *3*3*3*3
216=2*2*2*3*3*3
Б) НОК(350,400)=2*2*2*2*5*5*7=2800
350=2*5*5*7
400=2*2*2*2*5*5
В) НОК(60,220)=2*2*3*5*11=660
60=2*2*3*5
220=2*2*5*11
Г) НОК(23,47)=1081
Д) НОК(495,33)=495
495=5*3*3*11
33=3*11
Е) НОК(300,270) =2*2*3*3*3*5*5=2700
300=3*2*2*5*5
270=3*3*3*2*5
Ж) НОК(441, 4410).=4410
З) НОК(68,102,136)=2*2*2*3*17=408
68=2*2*17
102=3*2*17
136=2*2*2*17НОК (15,42,105) = ?
НОК (21,28,35) =?
НОК НАИМЕНЬШОЕ ОБЩЕЕ КРАТНОЕ
Решение: 1) 75=3*5*5
30=2*3*5
12=2*2*3
НОК (12,30,75)=2*2*3*5*5=300
2) 105=3*5*7
42=2*3*7
15=3*5
НОК (15,42,105)=2*3*5*7=210
3) 35=5*7
28=2*2*7
21=3*7
НОК (21,28,35)=3*2*2*5*7=420
1) 12 = 2*2*3
30 = 2*3*5
75 = 3*5*5
НОК = 2*2*3*5*5=300
2) 15 = 3*5
42 = 2*3*7
105 = 3*5*7
НОК = 2*3*5*7=210
3) 21 = 3*7
28 = 2*2*7
35 = 5*7
НОК = 2*2*3*5*7=420(21,28,35)=7
(15,42,105)=42 и 27Найди наименьшое общее кратное число:
а)18и45; б)30и40; в)210и350; г)20,70и15
Решение: а)18|2
9|3
3|3
1
45|3
15|3
5|5
1
2*3²*5=90
б)
30|2
15|3
5|5
1
40|2
20|2
10|2
5|5
1
2³*3*5=120
в)
210|2
105|3
35|5
7|7
1
350|2
175|5
35|5
7|7
1
2*3*5²*7=1050
г)
20|2
10|5
2|2
1
70|2
35|5
7|7
1
15|3
5|5
1
2²*3*5*7=420
Найдите наименьшое общее кратное число 504 и 540
Решение: Раскладываем каждое из чисел на простые множители
504 = 2·2·2·3·3·7
540 = 2·2·3·3·3·5
Теперь умножаем одно из чисел (например, 504) на множители, которые останутся в разложении второго числа (540) после вычеркивания множителей, общих для обоих чисел (выделено)
504·3·5 = 7560Найдите два натуральных числа, разность которых 66, а их наименьшее общее кратное равно 360.
Решение: Обозначим искомые числа через Х и У. Эти числа связаны соотношением У=Х+ 66. Ясно, что У>66. Число 360 делится на Х и на У.Тогда, 360= mX и 360=nY, где m и n натуральные числа. Возможные значения числа У находятся среди делителей числа 360 больших, чем 66.
Такими числами являются: 72, 90, 120, 180 и 360.
Соответствующие им значения числа Х: 6, 24, 54, 114 и 294. Числа 54, 114 и 294 не являются делителями числа 360. Следовательно, искомые значения числа Х находятся среди чисел: 6 и 24.
Пара чисел 6 и 72 не удовлетворяет условиям задачи, так как их НОК равен 72.
Пара 24 и 90 подходит.
ОТВЕТ: Одна пара чисел: 24 и 90.
Найдите два натуральных числа, разность которых 66, а их наименьшее общее кратное равно 360.
Решение: Обозначим искомые числа через Х и У. Эти числа связаны соотношением У=Х+ 66. Ясно, что У>66. Число 360 делится на Х и на У.Тогда, 360= mX и 360=nY, где m и n натуральные числа. Возможные значения числа У находятся среди делителей числа 360 больших, чем 66.Такими числами являются: 72, 90, 120, 180 и 360.Соответствующие им значения числа Х: 6, 24, 54, 114 и 294. Числа 54, 114 и 294 не являются делителями числа 360. Следовательно, искомые значения числа Х находятся среди чисел: 6 и 24.Пара чисел 6 и 72 не удовлетворяет условиям задачи, так как их НОК равен 72.Пара 24 и 90 подходит.ОТВЕТ: Одна пара чисел: 24 и 90.
 Обозначается одним из следующих способов: исходя из этого наименьшое общее кратное чисел 135 и 5 это 135
Обозначается одним из следующих способов: исходя из этого наименьшое общее кратное чисел 135 и 5 это 135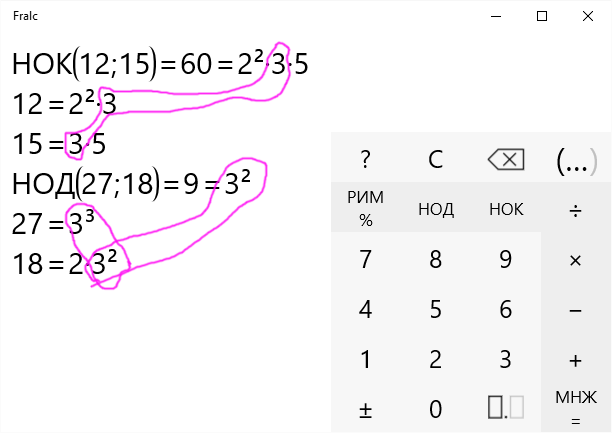
 ( НОК- наименьшее общее кратное) А) НОК(162,216). Б) НОК (350,400) В) НОК(60,220). Г) НОК(23,47) Д) НОК(495,33). Е) НОК(300,270) Ж) НОК(441, 4410). З) НОК(68,102,136)
( НОК- наименьшее общее кратное) А) НОК(162,216). Б) НОК (350,400) В) НОК(60,220). Г) НОК(23,47) Д) НОК(495,33). Е) НОК(300,270) Ж) НОК(441, 4410). З) НОК(68,102,136)

1 2 > >>
| 1 | Множитель | x^2-4 | |
| 2 | Множитель | 4x^2+20x+16 | |
| 3 | График | y=-x^2 | |
| 4 | Вычислить | 2+2 | |
| 5 | Множитель | x^2-25 | |
| 6 | Множитель | x^2+5x+6 | |
| 7 | Множитель | x^2-9 | |
| 8 | Множитель | x^3-8 | |
| 9 | Вычислить | квадратный корень из 12 | |
| 10 | Вычислить | квадратный корень из 20 | |
| 11 | Вычислить | квадратный корень из 50 | |
| 12 | Множитель | x^2-16 | |
| 13 | Вычислить | квадратный корень из 75 | |
| 14 | Множитель | x^2-1 | |
| 15 | Множитель | x^3+8 | |
| 16 | Вычислить | -2^2 | |
| 17 | Вычислить | квадратный корень из (-3)^4 | |
| 18 | Вычислить | квадратный корень из 45 | |
| 19 | Вычислить | квадратный корень из 32 | |
| 20 | Вычислить | квадратный корень из 18 | |
| 21 | Множитель | x^4-16 | |
| 22 | Вычислить | квадратный корень из 48 | |
| 23 | Вычислить | квадратный корень из 72 | |
| 24 | Вычислить | квадратный корень из (-2)^4 | |
| 25 | Множитель | x^3-27 | |
| 26 | Вычислить | -3^2 | |
| 27 | Множитель | x^4-1 | |
| 28 | Множитель | x^2+x-6 | |
| 29 | Множитель | x^3+27 | |
| 30 | Множитель | x^2-5x+6 | |
| 31 | Вычислить | квадратный корень из 24 | |
| 32 | Множитель | x^2-36 | |
| 33 | Множитель | x^2-4x+4 | |
| 34 | Вычислить | -4^2 | |
| 35 | Множитель | x^2-x-6 | |
| 36 | Множитель | x^4-81 | |
| 37 | Множитель | x^3-64 | |
| 38 | Вычислить | 4^3 | |
| 39 | Множитель | x^3-1 | |
| 40 | График | y=x^2 | |
| 41 | Вычислить | 2^3 | |
| 42 | Вычислить | (-12+ квадратный корень из -18)/60 | |
| 43 | Множитель | x^2-6x+9 | |
| 44 | Множитель | x^2-64 | |
| 45 | График | y=2x | |
| 46 | Множитель | x^3+64 | |
| 47 | Вычислить | (-8+ квадратный корень из -12)/40 | |
| 48 | Множитель | x^2-8x+16 | |
| 49 | Вычислить | 3^4 | |
| 50 | Вычислить | -5^2 | |
| 51 | Множитель | x^2-49 | |
| 52 | Вычислить | (-20+ квадратный корень из -75)/40 | |
| 53 | Множитель | x^2+6x+9 | |
| 54 | Множитель | 4x^2-25 | |
| 55 | Вычислить | квадратный корень из 28 | |
| 56 | Множитель | x^2-81 | |
| 57 | Вычислить | 2^5 | |
| 58 | Вычислить | -8^2 | |
| 59 | Вычислить | 2^4 | |
| 60 | Множитель | 4x^2-9 | |
| 61 | Вычислить | (-20+ квадратный корень из -50)/60 | |
| 62 | Вычислить | (-8+ квадратный корень из -20)/24 | |
| 63 | Множитель | x^2+4x+4 | |
| 64 | Множитель | x^2-10x+25 | |
| 65 | Вычислить | квадратный корень из -16 | |
| 66 | Множитель | x^2-2x+1 | |
| 67 | Вычислить | -7^2 | |
| 68 | График | f(x)=2^x | |
| 69 | Вычислить | 2^-2 | |
| 70 | Вычислить | квадратный корень из 27 | |
| 71 | Вычислить | квадратный корень из 80 | |
| 72 | Множитель | x^3+125 | |
| 73 | Вычислить | -9^2 | |
| 74 | Множитель | 2x^2-5x-3 | |
| 75 | Вычислить | квадратный корень из 40 | |
| 76 | Множитель | x^2+2x+1 | |
| 77 | Множитель | x^2+8x+16 | |
| 78 | График | y=3x | |
| 79 | Множитель | x^2+10x+25 | |
| 80 | Вычислить | 3^3 | |
| 81 | Вычислить | 5^-2 | |
| 82 | График | f(x)=x^2 | |
| 83 | Вычислить | квадратный корень из 54 | |
| 84 | Вычислить | (-12+ квадратный корень из -45)/24 | |
| 85 | Множитель | x^2+x-2 | |
| 86 | Вычислить | (-3)^3 | |
| 87 | Множитель | x^2-12x+36 | |
| 88 | Множитель | x^2+4 | |
| 89 | Вычислить | квадратный корень из (-8)^2 | |
| 90 | Множитель | x^2+7x+12 | |
| 91 | Вычислить | квадратный корень из -25 | |
| 92 | Множитель | x^2-x-20 | |
| 93 | Вычислить | 5^3 | |
| 94 | Множитель | x^2+8x+15 | |
| 95 | Множитель | x^2+7x+10 | |
| 96 | Множитель | 2x^2+5x-3 | |
| 97 | Вычислить квадратный корень | квадратный корень из 116 | |
| 98 | Множитель | x^2-x-12 | |
| 99 | Множитель | x^2-x-2 | |
| 100 | Вычислить | 2^2 |
Наименьшее общее кратное | Презентация к уроку по математике (5 класс):
Слайд 1
Наименьшее общее кратное чисел Тема урока :
Слайд 2
Подчеркнем общие делители пар чисел: 9: 18: 15: 20: 36: 48: 1, 3, 9 1, 2, 3, 6, 9, 18 1, 3, 5, 15 1, 2, 4, 5, 10, 20 1, 2, 3, 4, 6, 9, 12, 18, 36 1, 2, 3, 4, 6, 8, 12, 16, 42, 48 В каждой паре из общих делителей выделите наибольшее число:
Слайд 3
Наибольший Общий Делитель НОД Наибольший общий делитель чисел а и b или НОД(а, b ) –это наибольшее число, на которое делятся а , и b .
Слайд 4
— кратные 2 — кратные 3 : 2, 4, 6, 8, 10, 12, 14, 16, 18… : 3, 6, 9, 12, 15, 18, 21… Наименьшее общее кратное: 6 Подчеркнем общие кратные чисел: Из общих кратных выделим наименьшее число:
Слайд 5
Наименьшее Общее Кратное НОК Наименьшее общее кратное чисел а и b или НОК(а, b ) –это наименьшее число, которое делится и на а , и на b .
Слайд 6
Обозначение: НОК (2; 3) = 6. Алгоритм нахождения НОК: Пример: Найдите наименьшее общее кратное чисел: 75 и 60. 75 5 15 5 3 3 1 60 2 30 2 15 3 3 3 1 75 = 3 ∙ 5∙ 5 60 = 2∙ 2∙ 3 ∙ 5 НОК (75; 60) = 3 ∙ 5 ∙ 5∙ 2∙ 2 = 75∙ 2 ∙ 2 = 300. 1. Разложить все числа на простые множители. 2. Написать разложение одного из чисел (лучше наибольшего). 3. Дополнить данное разложение теми множителями из разложения других чисел, которые не вошли в написанное разложение. Определение. Наименьшее натуральное число, которое делится на каждое из данных натуральных чисел, называется наименьшим общим кратным.
Слайд 7
«Нахождение НОК и НОД натуральных чисел» Разложить числа на простые множители Если НОК (а; b) НОД (a;b) Найти и вычеркнуть одинаковые множители в одном из чисел (например b ). Выписать все множители числа a Дописать множители числа b , которые не вошли в состав разложения числа а Из всех разложений выбрать общие множители (У одного из чисел взять их в кружок) Выписать те числа, которые взяли в кружок Найти произведение полученных множителей
Выписать все множители числа a Дописать множители числа b , которые не вошли в состав разложения числа а Из всех разложений выбрать общие множители (У одного из чисел взять их в кружок) Выписать те числа, которые взяли в кружок Найти произведение полученных множителей
Слайд 8
Слово «крат» — старинное русское слово ( XI век), означающее «раз». Слова «многократно» означает «много раз». Понятием кратного пользуются в жизненной практике при установлении вида года. Через каждые три обыкновенных года, в каждом из которых по 365 дней (в феврале 28 дней), бывает четвертый год, так называемый високосный, в котором 366 дней (в феврале 29 дней). Если число, которым выражается указанный год, есть число, кратное 4, то указанный год високосный, а если не кратно 4, то год обыкновенный. Так, 2008 год — високосный, так как 2008 кратно 4, 2007 — не високосный, так как 2007 не кратно 4. Историческая минутка. 8
Слайд 9
НОД (60,75) = ? 60 30 15 5 1 2 2 3 5 75 25 5 1 3 5 5 НОД (60,75) = 3 · 5 = 15
Слайд 10
б) НОК (12; 16) = 2 ∙ 2 ∙ 2 ∙ 2 ∙ 3 = 48; Найдите наименьшее общее кратное чисел: б)12 и 16; I 12 2 6 2 3 3 1 16 2 8 2 4 2 2 2 1 12 = 2 ∙ 2 ∙ 3; 16 = 2 ∙ 2 ∙ 2 ∙ 2
Слайд 11
Найдите наименьшее общее кратное чисел: г) 396 и 180; г) НОК (396; 180) = 2 ∙ 2 ∙ 3 ∙ 3 ∙ 11 ∙ 5 = 1980; II 396 2 198 2 99 3 33 3 11 11 1 180 2 90 2 45 3 15 3 5 5 1 396 = 2 ∙ 2 ∙ 3 ∙ 3 · 11 ; 18 0 = 2 ∙ 2 ∙ 3 ∙ 3 · 5
Слайд 12
е) 168, 231 и 60. Найдите наименьшее общее кратное чисел: е) НОК (168; 231; 60) = 231 ∙ 2 ∙ 2 ∙ 2 ∙ 3 · 7 · 5 = 194 040 III 168 2 84 2 42 2 21 3 7 7 1 60 2 30 2 15 3 5 5 1 231 231 1 168 = 2 ∙ 2 ∙ 2 ∙ 3 · 7 ; 231 = 1 · 231 ; 60 = 2 ∙ 2 ∙ 3 ∙ 5
Найдите наименьшее общее кратное чисел: е) НОК (168; 231; 60) = 231 ∙ 2 ∙ 2 ∙ 2 ∙ 3 · 7 · 5 = 194 040 III 168 2 84 2 42 2 21 3 7 7 1 60 2 30 2 15 3 5 5 1 231 231 1 168 = 2 ∙ 2 ∙ 2 ∙ 3 · 7 ; 231 = 1 · 231 ; 60 = 2 ∙ 2 ∙ 3 ∙ 5
Слайд 13
Являются ли числа 54 и 65 взаимно простыми? Найдите наименьшее общее кратное чисел 54 и 65. Равно ли оно произведению 54 и 65? Запишите какие-нибудь два взаимно простых числа. Найдите наименьшее общее кратное этих чисел. Сделайте вывод. 54 2 27 3 9 3 3 3 1 65 5 13 13 1 54 = 2 ∙ 3 ∙ 3 ∙ 3 65 = 5 ∙ 13 НОК (54; 65) = 54 · 65 = 3510; 24 2 12 2 6 2 3 3 1 35 5 7 7 1 24 = 2 ∙ 2 ∙ 2 ∙ 3 35 = 5 · 7 НОК (24; 35) = 24 · 35 = 840; Вывод: Наименьшее общее кратное двух взаимно простых чисел равно их произведению. Найдем наименьшее общее кратное любых двух взаимно простых чисел
Слайд 14
Найдите наименьшее общее кратное чисел: а) 45 и 135; б) 34 и 170. Равно ли оно одному из данных чисел? Так как большее число делится на меньшее, то наименьшим общим кратным этих чисел будет являться большее число. НОК (45; 135) = 135; НОК (34; 170) = 170. Наибольший общий делитель этих чисел: НОД (45; 135) = 45; НОД (34; 170) = 34. 45 3 15 3 5 5 1 135 3 45 3 15 3 5 5 1 34 2 17 17 1 170 2 85 5 17 17 1
НОК (45; 135) = 135; НОК (34; 170) = 170. Наибольший общий делитель этих чисел: НОД (45; 135) = 45; НОД (34; 170) = 34. 45 3 15 3 5 5 1 135 3 45 3 15 3 5 5 1 34 2 17 17 1 170 2 85 5 17 17 1
Слайд 15
НОД (22; 66) = 2 · 11 = 22 Найдите наименьшее общее кратное и наибольший общий делитель чисел 22 и 66: в) НОК (22; 66) = ? НОД (22; 66) = ? 22 2 11 11 1 66 2 33 3 11 11 1 НОК (22; 66) = 2 · 3 · 11 = 66 I
Слайд 16
НОД (39; 65) = 13. Найдите наименьшее общее кратное и наибольший общий делитель чисел 39 и 65: г) НОК (39; 65) = ? НОД (39; 65) = ? 39 3 13 13 1 65 5 13 13 1 НОК (39; 65) = 5 · 13 · 3 = 195; II
Слайд 17
Ответить на вопросы: Какое число называют наименьшим общим кратным натуральных чисел а и b ? Какое число называют наибольшим общим делителем натуральных чисел а и b ? Какое число является наименьшим общим кратным чисел т и п, если число т кратно числу n ? Какие натуральные числа называют простыми? Какие натуральные числа называют взаимно простыми? 10. 05.2012 17 www.konspekturoka.ru
05.2012 17 www.konspekturoka.ru
Слайд 18
НОК (60,75) = ? 60 30 15 5 1 2 2 3 5 75 25 5 1 3 5 5 НОК (60,75) = 2 · 2 · 3 · 5 = 300 · 5 60 · 5 = 300 НОК (60,75) =
Слайд 19
? 60 30 15 5 1 2 2 3 5 42 21 7 1 2 3 7 НОД (60,42) = 2 = 6 · 3 60 · 7 = 420 НОК (60,42) =
Слайд 20
НОК (250; 130)= ? 250 50 10 2 1 5 5 5 2 130 10 2 1 13 5 2 НОД (250;130)= 5 · 2 = 10 250 · 13 = 3250
Слайд 21
НОД (36,48)= ? 36 18 9 3 1 2 2 3 3 48 24 12 6 3 1 2 2 2 2 3 НОД (36,48)= 2 · 2 · 3 = 12
Слайд 22
НОК (36, 48) = ? 36 18 9 3 1 2 2 3 3 48 24 12 6 3 1 2 2 2 2 3 НОК (36, 48) = 36 · =144 2 · 2
Слайд 23
НОК (360;432)= ? 360 120 40 8 4 2 1 3 3 5 2 2 2 432 144 48 24 12 6 3 1 3 3 2 2 2 2 3 НОД (360;432)= 3 · 3 · 2 · 2 · 2 = 72 360 2160 · 2 · 3 =
Слайд 24
ЗАДАНИЕ НА ДОМ: №664
Слайд 25
СПАСИБО ЗА УРОК!
| 1 | Оценка с использованием заданного значения | квадратный корень из 50 | |
| 2 | Оценка с использованием заданного значения | квадратный корень из 45 | |
| 3 | Оценить | 5+5 | |
| 4 | Оценить | 7*7 | |
| 5 | Найти простую факторизацию | 24 | |
| 6 | Преобразование в смешанный номер | 52/6 | |
| 7 | Преобразование в смешанный номер | 93/8 | |
| 8 | Преобразование в смешанный номер | 34/5 | |
| 9 | График | у=х+1 | |
| 10 | Оценка с использованием заданного значения | квадратный корень из 128 | |
| 11 | Найдите площадь поверхности | сфера (3) | |
| 12 | Оценить | 54-6÷2+6 | |
| 13 | График | г=-2x | |
| 14 | Оценить | 8*8 | |
| 15 | Преобразование в десятичное число | 5/9 | |
| 16 | Оценка с использованием заданного значения | квадратный корень из 180 | |
| 17 | График | у=2 | |
| 18 | Преобразование в смешанный номер | 7/8 | |
| 19 | Оценить | 9*9 | |
| 20 | Решите для C | С=5/9*(Ф-32) | |
| 21 | Упростить | 1/3+1 1/12 | |
| 22 | График | у=х+4 | |
| 23 | График | г=-3 | |
| 24 | График | х+у=3 | |
| 25 | График | х=5 | |
| 26 | Оценить | 6*6 | |
| 27 | Оценить | 2*2 | |
| 28 | Оценить | 4*4 | |
| 29 | Оценить | 1/2+(2/3)÷(3/4)-(4/5*5/6) | |
| 30 | Оценить | 1/3+13/12 | |
| 31 | Оценка | 5*5 | |
| 32 | Решить для d | 2д=5в(о)-вр | |
| 33 | Преобразование в смешанный номер | 3/7 | |
| 34 | График | г=-2 | |
| 35 | Найдите склон | ||
| 36 | Преобразование в проценты | 9 | |
| 37 | График | у=2х+2 | |
| 38 | 92+5х+6=0|||
| 41 | Преобразование в смешанный номер | 1/6 | |
| 42 | Преобразование в десятичное число | 9% | |
| 43 | Найти n | 12н-24=14н+28 | |
| 44 | Оценить | 16*4 | |
| 45 | Упростить | кубический корень из 125 | |
| 46 | Преобразование в упрощенную дробь | 43% | |
| 47 | График | х=1 | |
| 48 | График | у=6 | |
| 49 | График | г=-7 | |
| 50 | График | у=4х+2 | |
| 51 | Найдите склон | у=7 | |
| 52 | у=3х+4 | ||
| 53 | График | у=х+5 | |
| 54 | График | 92-9=0||
| 58 | Оценка с использованием заданного значения | квадратный корень из 192 | |
| 59 | Оценка с использованием заданного значения | квадратный корень из 25/36 | |
| 60 | Найти простую факторизацию | 14 | |
| 61 | Преобразование в смешанный номер | 7/10 | |
| 62 | Решите для | (-5а)/2=75 | |
| 63 | Упростить | х | |
| 64 | Оценить | 6*4 | |
| 65 | Оценить | 6+6 | |
| 66 | Оценить | -3-5 | |
| 67 | Оценить | -2-2 | |
| 68 | Упростить | квадратный корень из 1 | |
| 69 | квадратный корень из 4 | ||
| 70 | Найди обратное | 1/3 | |
| 71 | Преобразование в смешанный номер | 20. 11. 11. | |
| 72 | Преобразование в смешанный номер | 7/9 | |
| 73 | Найти LCM | 11, 13, 5, 15, 14 | , , , , |
| 76 | График | 3x+4y=12 | |
| 77 | График | 3x-2y=6 | |
| 78 | График | у=-х-2 | |
| 79 | График | у=3х+7 | |
| 80 | Определить, является ли многочлен | 2x+2 | |
| 81 | График | у=2х-6 | |
| 82 | График | у=2х-7 | |
| 83 | График | у=2х-2 | |
| 84 | График | у=-2х+1 | |
| 85 | График | у=-3х+4 | |
| 86 | График | у=-3х+2 | |
| 87 | График | у=х-4 | |
| 88 | Оценить | (4/3)÷(7/2) | |
| 89 | График | 2x-3y=6 | |
| 90 | График | х+2у=4 | |
| 91 | График | х=7 | |
| 92 | График | х-у=5 | |
| 93 | Решение с использованием свойства квадратного корня 92-2x-3=0 | ||
| 95 | Найдите площадь поверхности | конус (12)(9) | |
| 96 | Преобразование в смешанный номер | 3/10 | |
| 97 | Преобразование в смешанный номер | 7/20 | 92)
LCM 15, 27 и 45
Калькуляторы Учебные ресурсы по математике
- Домашняя страница
- Математические функции
- Калькулятор НОК
- LCM 15, 27 и 45
LCM 15, 27 и 45 равно 135. Всесторонняя работа дает представление о том, как найти, что больше 135. это lcm 15, 27 и 45 с использованием простых множителей и специальных методов деления, а также пример использования математики и реальных задач.
Всесторонняя работа дает представление о том, как найти, что больше 135. это lcm 15, 27 и 45 с использованием простых множителей и специальных методов деления, а также пример использования математики и реальных задач.
какой lcm у 15, 27 и 45?
lcm (15 27 45) = (?)
15 => 3 x 5
27 => 3 x 3 x 3
45 => 3 x 3 x 5
= 3 x 3 x 5 x 3
= 135
lcm (15, 27 и 45) = 135
135 lcm единиц 15, 27 и 45. из 15, 27 и 45,
{3, 3, 5} в {3 x 5, 3 x 3 x 3, 3 x 3 x 5} являются наиболее повторяющимися факторами 15, 27 и 45,
{3} в {3 x 5, 3 x 3 x 3, 3 x 3 x 5} – другие оставшиеся множители чисел 15, 27 и 45.
Использование в математике: НОК 15, 27 и 45
Ниже приведены некоторые математические приложения, в которых можно использовать lcm 15, 27 и 45:
- найти наименьшее число, которое точно делится на 15, 27 и 45.
- , чтобы найти общие знаменатели для дробей, имеющих 15, 27 и 45 в качестве знаменателей при сложении или вычитании разнородных дробей.

Использование в реальных задачах: 15, 27 и 45 lcm
разное расписание происходит вместе в одно и то же время. Например, задачи реального мира включают lcm в ситуации, когда нужно определить, в какое время все колокола A, B и C звонят вместе, если колокол A звонит через 15 секунд, B звонит через 27 секунд и C многократно звонит через 45 секунд. Ответ заключается в том, что все колокола A, B и C звонят вместе через 135 секунд в первый раз, через 270 секунд во второй раз, через 405 секунд в третий раз и так далее.
Важные примечания: 15, 27 и 45 lcm
Ниже приведены важные примечания, которые следует помнить при решении lcm из 15, 27 и 45:
- Повторяющиеся и неповторяющиеся простые множители 15, 27 и 45 следует умножить, чтобы найти наименьшее общее кратное 15, 27 и 45, при решении lcm с использованием метода простых множителей.
- Результаты lcm 15, 27 и 45 идентичны, даже если мы изменим порядок заданных чисел в вычислении lcm, это означает, что порядок заданных чисел в вычислении lcm не повлияет на результаты.

Для значений, отличных от 15, 27 и 45, используйте этот инструмент ниже:
В приведенном ниже решенном примере с пошаговой работой показано, как найти lcm числа 15, 27 и 45, используя либо метод простых множителей, либо метод специального деления. .
Пример решения с использованием метода простых множителей:
Чему равно НОК 15, 27 и 45?
шаг 1
Обратитесь к входным параметрам, значениям и посмотрите, что будет найдено:
Входные параметры и значения:
A = 15
B = 27
C = 45
Что нужно найти:
найдите lcm числа 15, 27 и 45
шаг 2 Найдите простые множители числа 1
Простые множители 15 = 3 x 5
Простые множители 27 = 3 x 3 x 3
Простые множители 45 = 3 x 3 x 5
шаг 3 Определите повторяющиеся и неповторяющиеся простые делители чисел 15, 27 и 45 :
{3, 3, 5} — наиболее повторяющиеся факторы, а {3} — неповторяющиеся факторы 15, 27 и 45.
шаг 4 Найдите произведение повторяющихся и неповторяющихся простых множителей 15, 27 и 45:
= 3 x 3 x 5 x 3
= 135
lcm(20 и 30) = 135
Следовательно,
lcm 15, 27 и 45 равно 135
Пример решения с использованием специального метода деления:
Этот специальный метод деления является самым простым способом понять весь расчет того, что такое lcm для 15, 27 и 45.
Шаг 1 Адресуйте входные параметры, значения и наблюдайте, что нужно найти:
Входные параметры и значения:
Целые числа: 15, 27 и 45
Что нужно найти:
lcm (15, 27, 45) = ?
шаг 2 Расположите заданные целые числа по горизонтали, разделяя их пробелами или запятыми. Формат:
15, 27 и 45
шаг 3 Выберите делитель, который делит каждое или большинство заданных целых чисел (15, 27 и 45), разделите каждое целое число отдельно и запишите частное в следующей строке прямо под соответствующими целыми числами. Перенесите целое число на следующую строку, если какое-либо целое число в числах 15, 27 и 45 не делится на выбранный делитель; повторяйте тот же процесс, пока все целые числа не будут равны 1, как показано ниже:
Перенесите целое число на следующую строку, если какое-либо целое число в числах 15, 27 и 45 не делится на выбранный делитель; повторяйте тот же процесс, пока все целые числа не будут равны 1, как показано ниже:
| 3 | 15 | 27 | 45 |
| 3 | 5 | 9 | 15 |
| 3 | 5 | 3 | 5 |
| 5 | 5 | 1 | 5 |
| 1 | 1 | 1 |
Шаг 4 Умножение дивизоров, чтобы найти LCM 15, 27 и 45:
= 3 x 3 x 5
. = 135
LCM(15, 27, 45) = 135
Наименьшее общее кратное для трех чисел 15, 27 и 45 равно 135
Наименьшее общее кратное — 135
| Числа | ЛКМ |
|---|---|
| 135 и 1 | 135 |
| 135 и 2 | 270 |
| 135 и 3 | 135 |
| 135 и 4 | 540 |
| 135 и 5 | 135 |
| 135 и 6 | 270 |
| 135 и 7 | 945 |
| 135 и 8 | 1080 |
| 135 и 9 | 135 |
| 135 и 10 | 270 |
| 135 и 11 | 1 485 |
| 135 и 12 | 540 |
| 135 и 13 | 1 755 |
| 135 и 14 | 1 890 |
| 135 и 15 | 135 |
| 135 и 16 | 2 160 |
| 135 и 17 | 2 295 |
| 135 и 18 | 270 |
| 135 и 19 | 2 565 |
| 135 и 20 | 540 |
| 135 и 21 | 945 |
| 135 и 22 | 2 970 |
| 135 и 23 | 3 105 |
| 135 и 24 | 1080 |
| 135 и 25 | 675 |
| 135 и 26 | 3 510 |
| 135 и 27 | 135 |
| 135 и 28 | 3 780 |
| 135 и 29 | 3 915 |
| 135 и 30 | 270 |
| 135 и 31 | 4 185 |
| 135 и 32 | 4 320 |
| 135 и 33 | 1 485 |
| 135 и 34 | 4 590 |
| 135 и 35 | 945 |
| 135 и 36 | 540 |
| 135 и 37 | 4 995 |
| 135 и 38 | 5 130 |
| 135 и 39 | 1 755 |
| 135 и 40 | 1 080 |
| 135 и 41 | 5 535 |
| 135 и 42 | 1 890 |
| 135 и 43 | 5 805 |
| 135 и 44 | 5 940 |
| 135 и 45 | 135 |
| 135 и 46 | 6 210 |
| 135 и 47 | 6 345 |
| 135 и 48 | 2 160 |
| 135 и 49 | 6 615 |
| 135 и 50 | 1 350 |
| 135 и 51 | 2 295 |
| 135 и 52 | 7 020 |
| 135 и 53 | 7 155 |
| 135 и 54 | 270 |
| 135 и 55 | 1 485 |
| 135 и 56 | 7 560 |
| 135 и 57 | 2 565 |
| 135 и 58 | 7 830 |
| 135 и 59 | 7 965 |
| 135 и 60 | 540 |
| 135 и 61 | 8 235 |
| 135 и 62 | 8 370 |
| 135 и 63 | 945 |
| 135 и 64 | 8 640 |
| 135 и 65 | 1 755 |
| 135 и 66 | 2 970 |
| 135 и 67 | 9045 |
| 135 и 68 | 9 180 |
| 135 и 69 | 3 105 |
| 135 и 70 | 1 890 |
| 135 и 71 | 9 585 |
| 135 и 72 | 1080 |
| 135 и 73 | 9 855 |
| 135 и 74 | 9 990 |
| 135 и 75 | 675 |
| Номера | ЛКМ |
|---|---|
| 135 и 76 | 10 260 |
| 135 и 77 | 10 395 |
| 135 и 78 | 3 510 |
| 135 и 79 | 10 665 |
| 135 и 80 | 2 160 |
| 135 и 81 | 405 |
| 135 и 82 | 11 070 |
| 135 и 83 | 11 205 |
| 135 и 84 | 3 780 |
| 135 и 85 | 2 295 |
| 135 и 86 | 11 610 |
| 135 и 87 | 3 915 |
| 135 и 88 | 11 880 |
| 135 и 89 | 12 015 |
| 135 и 90 | 270 |
| 135 и 91 | 12 285 |
| 135 и 92 | 12 420 |
| 135 и 93 | 4 185 |
| 135 и 94 | 12 690 |
| 135 и 95 | 2 565 |
| 135 и 96 | 4 320 |
| 135 и 97 | 13 095 |
| 135 и 98 | 13 230 |
| 135 и 99 | 1 485 |
| 135 и 100 | 2 700 |
| 135 и 101 | 13 635 |
| 135 и 102 | 4 590 |
| 135 и 103 | 13 905 |
| 135 и 104 | 14 040 |
| 135 и 105 | 945 |
| 135 и 106 | 14 310 |
| 135 и 107 | 14 445 |
| 135 и 108 | 540 |
| 135 и 109 | 14 715 |
| 135 и 110 | 2 970 |
| 135 и 111 | 4 995 |
| 135 и 112 | 15 120 |
| 135 и 113 | 15 255 |
| 135 и 114 | 5 130 |
| 135 и 115 | 3 105 |
| 135 и 116 | 15 660 |
| 135 и 117 | 1 755 |
| 135 и 118 | 15 930 |
| 135 и 119 | 16 065 |
| 135 и 120 | 1080 |
| 135 и 121 | 16 335 |
| 135 и 122 | 16 470 |
| 135 и 123 | 5 535 |
| 135 и 124 | 16 740 |
| 135 и 125 | 3 375 |
| 135 и 126 | 1 890 |
| 135 и 127 | 17 145 |
| 135 и 128 | 17 280 |
| 135 и 129 | 5 805 |
| 135 и 130 | 3 510 |
| 135 и 131 | 17 685 |
| 135 и 132 | 5 940 |
| 135 и 133 | 17 955 |
| 135 и 134 | 18 090 |
| 135 и 135 | 135 |
| 135 и 136 | 18 360 |
| 135 и 137 | 18 495 |
| 135 и 138 | 6 210 |
| 135 и 139 | 18 765 |
| 135 и 140 | 3 780 |
| 135 и 141 | 6 345 |
| 135 и 142 | 19 170 |
| 135 и 143 | 19 305 |
| 135 и 144 | 2 160 |
| 135 и 145 | 3 915 |
| 135 и 146 | 19 710 |
| 135 и 147 | 6 615 |
| 135 и 148 | 19 980 |
| 135 и 149 | 20 115 |
| 135 и 150 | 1 350 |
| Номера | ЛКМ |
|---|---|
| 135 и 151 | 20 385 |
| 135 и 152 | 20 520 |
| 135 и 153 | 2 295 |
| 135 и 154 | 20 790 |
| 135 и 155 | 4 185 |
| 135 и 156 | 7 020 |
| 135 и 157 | 21 195 |
| 135 и 158 | 21 330 |
| 135 и 159 | 7 155 |
| 135 и 160 | 4 320 |
| 135 и 161 | 21 735 |
| 135 и 162 | 810 |
| 135 и 163 | 22 005 |
| 135 и 164 | 22 140 |
| 135 и 165 | 1 485 |
| 135 и 166 | 22 410 |
| 135 и 167 | 22 545 |
| 135 и 168 | 7 560 |
| 135 и 169 | 22 815 |
| 135 и 170 | 4 590 |
| 135 и 171 | 2 565 |
| 135 и 172 | 23 220 |
| 135 и 173 | 23 355 |
| 135 и 174 | 7 830 |
| 135 и 175 | 4 725 |
| 135 и 176 | 23 760 |
| 135 и 177 | 7 965 |
| 135 и 178 | 24 030 |
| 135 и 179 | 24 165 |
| 135 и 180 | 540 |
| 135 и 181 | 24 435 |
| 135 и 182 | 24 570 |
| 135 и 183 | 8 235 |
| 135 и 184 | 24 840 |
| 135 и 185 | 4 995 |
| 135 и 186 | 8 370 |
| 135 и 187 | 25 245 |
| 135 и 188 | 25 380 |
| 135 и 189 | 945 |
| 135 и 190 | 5 130 |
| 135 и 191 | 25 785 |
| 135 и 192 | 8 640 |
| 135 и 193 | 26 055 |
| 135 и 194 | 26 190 |
| 135 и 195 | 1 755 |
| 135 и 196 | 26 460 |
| 135 и 197 | 26 595 |
| 135 и 198 | 2 970 |
| 135 и 199 | 26 865 |
| 135 и 200 | 5 400 |
| 135 и 201 | 9045 |
| 135 и 202 | 27 270 |
| 135 и 203 | 27 405 |
| 135 и 204 | 9 180 |
| 135 и 205 | 5 535 |
| 135 и 206 | 27 810 |
| 135 и 207 | 3 105 |
| 135 и 208 | 28 080 |
| 135 и 209 | 28 215 |
| 135 и 210 | 1 890 |
| 135 и 211 | 28 485 |
| 135 и 212 | 28 620 |
| 135 и 213 | 9 585 |
| 135 и 214 | 28 890 |
| 135 и 215 | 5 805 |
| 135 и 216 | 1 080 |
| 135 и 217 | 29 295 |
| 135 и 218 | 29 430 |
| 135 и 219 | 9 855 |
| 135 и 220 | 5 940 |
| 135 и 221 | 29 835 |
| 135 и 222 | 9 990 |
| 135 и 223 | 30 105 |
| 135 и 224 | 30 240 |
| 135 и 225 | 675 |
| Номера | ЛКМ |
|---|---|
| 135 и 226 | 30 510 |
| 135 и 227 | 30 645 |
| 135 и 228 | 10 260 |
| 135 и 229 | 30 915 |
| 135 и 230 | 6 210 |
| 135 и 231 | 10 395 |
| 135 и 232 | 31 320 |
| 135 и 233 | 31 455 |
| 135 и 234 | 3 510 |
| 135 и 235 | 6 345 |
| 135 и 236 | 31 860 |
| 135 и 237 | 10 665 |
| 135 и 238 | 32 130 |
| 135 и 239 | 32 265 |
| 135 и 240 | 2 160 |
| 135 и 241 | 32 535 |
| 135 и 242 | 32 670 |
| 135 и 243 | 1 215 |
| 135 и 244 | 32 940 |
| 135 и 245 | 6 615 |
| 135 и 246 | 11 070 |
| 135 и 247 | 33 345 |
| 135 и 248 | 33 480 |
| 135 и 249 | 11 205 |
| 135 и 250 | 6 750 |
| 135 и 251 | 33 885 |
| 135 и 252 | 3 780 |
| 135 и 253 | 34 155 |
| 135 и 254 | 34 290 |
| 135 и 255 | 2 295 |
| 135 и 256 | 34 560 |
| 135 и 257 | 34 695 |
| 135 и 258 | 11 610 |
| 135 и 259 | 34 965 |
| 135 и 260 | 7 020 |
| 135 и 261 | 3 915 |
| 135 и 262 | 35 370 |
| 135 и 263 | 35 505 |
| 135 и 264 | 11 880 |
| 135 и 265 | 7 155 |
| 135 и 266 | 35 910 |
| 135 и 267 | 12 015 |
| 135 и 268 | 36 180 |
| 135 и 269 | 36 315 |
| 135 и 270 | 270 |
| 135 и 271 | 36 585 |
| 135 и 272 | 36 720 |
| 135 и 273 | 12 285 |
| 135 и 274 | 36 990 |
| 135 и 275 | 7 425 |
| 135 и 276 | 12 420 |
| 135 и 277 | 37 395 |
| 135 и 278 | 37 530 |
| 135 и 279 | 4 185 |
| 135 и 280 | 7 560 |
| 135 и 281 | 37 935 |
| 135 и 282 | 12 690 |
| 135 и 283 | 38 205 |
| 135 и 284 | 38 340 |
| 135 и 285 | 2 565 |
| 135 и 286 | 38 610 |
| 135 и 287 | 38 745 |
| 135 и 288 | 4 320 |
| 135 и 289 | 39 015 |
| 135 и 290 | 7 830 |
| 135 и 291 | 13 095 |
| 135 и 292 | 39 420 |
| 135 и 293 | 39 555 |
| 135 и 294 | 13 230 |
| 135 и 295 | 7 965 |
| 135 и 296 | 39 960 |
| 135 и 297 | 1 485 |
| 135 и 298 | 40 230 |
| 135 и 299 | 40 365 |
| 135 и 300 | 2,700 |
Drepmel — ресурс по перепрофилированию лекарств от меланомы Multi-Omics для определения приоритетности комбинаций препаратов и понимания микроокружения опухоли
Клетки. 2022 сен; 11 (18): 2894.
2022 сен; 11 (18): 2894.
Опубликовано в Интернете 2022 г. , 3 , 4 , 2 , 5 и 2, *
Эсмаил Эбрахими, академический редактор, Матиас Коль, академический редактор, и Джеймс Перкинс, академический редактор, информация о лицензии
9091 информация Заявление об отказе от ответственности- Дополнительные материалы
- Заявление о доступности данных
Несмотря на то, что был достигнут значительный прогресс в лечении пациентов с запущенной меланомой с помощью таргетной и иммунотерапии, de novo и приобретенная резистентность являются обычным явлением. После неэффективности лечения терапевтические возможности очень ограничены, и срочно необходимы новые стратегии. Комбинированная терапия часто более эффективна, чем отдельные препараты, и в настоящее время широко используется в клинической практике. Таким образом, существует острая потребность во всеобъемлющем вычислительном ресурсе для определения рациональной комбинированной терапии. Мы разработали приложение Shiny, DRepMel, чтобы обеспечить рациональные прогнозы комбинированного лечения для пациентов с меланомой из семидесяти трех тысяч комбинаций на основе многокомпонентного вычислительного подхода с перепрофилированием лекарств с использованием полного секвенирования экзома и данных секвенирования РНК в массовых образцах из двух независимых когорт пациентов. DRepMel предоставляет надежные прогнозы в качестве ресурса, а также определяет потенциальное воздействие лечения на микроокружение опухоли (TME) с использованием данных одноклеточной РНК-секвенации пациентов с меланомой. Доступность: DRepMel доступен онлайн.
Мы разработали приложение Shiny, DRepMel, чтобы обеспечить рациональные прогнозы комбинированного лечения для пациентов с меланомой из семидесяти трех тысяч комбинаций на основе многокомпонентного вычислительного подхода с перепрофилированием лекарств с использованием полного секвенирования экзома и данных секвенирования РНК в массовых образцах из двух независимых когорт пациентов. DRepMel предоставляет надежные прогнозы в качестве ресурса, а также определяет потенциальное воздействие лечения на микроокружение опухоли (TME) с использованием данных одноклеточной РНК-секвенации пациентов с меланомой. Доступность: DRepMel доступен онлайн.
Ключевые слова: мультиомика, меланома, перепрофилирование лекарств, микроокружение
Меланома кожи — наиболее опасная форма рака кожи, склонная к агрессивному метастазированию во многие органы [1]. Меланома долгое время была образцом персонализированной медицины с таргетной терапией, такой как ингибиторы BRAF и комбинация ингибиторов BRAF-MEK, которые высокоэффективны против 50% меланом с активирующими мутациями BRAF. Несмотря на это, отсутствуют таргетные методы лечения меланомы с мутацией NRAS и примерно 25% меланом, у которых не выявлены онкогенные драйверные мутации. В последнее время иммунотерапия, такая как анти-PD1 и анти-CTLA4, показала большие перспективы для улучшения результатов лечения пациентов. Однако после неэффективности лечения доступны ограниченные варианты лечения. Были разработаны и одобрены комбинации лекарств для преодоления резистентности к таргетной и иммунотерапии, однако вычислительные методы и ресурсы были ограничены для прогнозирования комбинаций лекарств. Здесь мы разработали три мультиомных подхода к прогнозированию эффективности комбинированной терапии с использованием двух независимых когорт данных о пациентах с меланомой, а результаты отображаются в удобном для пользователя приложении Shiny DRepMel. Было показано, что подходы машинного обучения и глубокого обучения эффективны для прогнозирования комбинированной терапии рака с использованием данных о линиях раковых клеток [2,3,4].
Несмотря на это, отсутствуют таргетные методы лечения меланомы с мутацией NRAS и примерно 25% меланом, у которых не выявлены онкогенные драйверные мутации. В последнее время иммунотерапия, такая как анти-PD1 и анти-CTLA4, показала большие перспективы для улучшения результатов лечения пациентов. Однако после неэффективности лечения доступны ограниченные варианты лечения. Были разработаны и одобрены комбинации лекарств для преодоления резистентности к таргетной и иммунотерапии, однако вычислительные методы и ресурсы были ограничены для прогнозирования комбинаций лекарств. Здесь мы разработали три мультиомных подхода к прогнозированию эффективности комбинированной терапии с использованием двух независимых когорт данных о пациентах с меланомой, а результаты отображаются в удобном для пользователя приложении Shiny DRepMel. Было показано, что подходы машинного обучения и глубокого обучения эффективны для прогнозирования комбинированной терапии рака с использованием данных о линиях раковых клеток [2,3,4]. Чтобы оценить потенциальные лечебные эффекты таргетной и иммунотерапии, мы использовали данные omics от пациентов с меланомой, которые состоят как из опухолевых, так и из иммунных / стромальных клеток, а не из наборов данных о линиях раковых клеток. Кроме того, в приложение было включено целевое иммунное микроокружение опухоли (TME), полученное из данных scRNA-seq образцов пациентов с меланомой [5], для понимания потенциального влияния комбинированной терапии на TME.
Чтобы оценить потенциальные лечебные эффекты таргетной и иммунотерапии, мы использовали данные omics от пациентов с меланомой, которые состоят как из опухолевых, так и из иммунных / стромальных клеток, а не из наборов данных о линиях раковых клеток. Кроме того, в приложение было включено целевое иммунное микроокружение опухоли (TME), полученное из данных scRNA-seq образцов пациентов с меланомой [5], для понимания потенциального влияния комбинированной терапии на TME.
Мы разработали комплексный подход к перепрофилированию лекарств и прогнозированию комбинированной терапии для пациентов с меланомой и применили его к двум независимым когортам пациентов с меланомой с соответствующими наборами данных Whole Exome Sequence (WES) и RNA-seq от одних и тех же пациентов: TCGA (N = 459 ) и когорты меланомы Моффита (N = 135; ). Когорта меланомы Моффита (N = 135) была описана в нашей предыдущей работе [6]. Вкратце, это исследование (MCC# 19147) проводилось в соответствии с признанными этическими принципами (например, Хельсинкская декларация, CIOMS, Белмонтский отчет, Общее правило США) и было одобрено Чесапикским институциональным наблюдательным советом (IRB). Отказ от согласия был предоставлен Chesapeake IRB. Мы включаем дополнительную информацию об анализе последовательности всего экзома (WES) и анализе последовательности РНК здесь. Сводная информация включена в .
Отказ от согласия был предоставлен Chesapeake IRB. Мы включаем дополнительную информацию об анализе последовательности всего экзома (WES) и анализе последовательности РНК здесь. Сводная информация включена в .
Открыть в отдельном окне
Рабочий процесс DRepMel: интегративный многокомпонентный подход к перепрофилированию лекарств для прогнозирования комбинированной терапии пациентов с меланомой с использованием двух независимых когорт пациентов с меланомой (с соответствующими наборами данных Whole Exome Sequence (WES) и RNA-seq из те же пациенты: когорты TCGA (N = 459) и меланомы Моффита (N = 135)).
Таблица 1
Информация о размере выборки, доступных метаданных и секвенировании ДНК/РНК когорт TCGA и Moffitt.
| TCGA (N = 459) | Moffitt (N = 135) | ||
|---|---|---|---|
| Когорты мутаций n (%) | |||
| BRAF | 236 (51,4) | 59 (43,7) | |
| NRAS | 125 (27,2) | 34 (25,2) | |
| Triple Wild | 88 (19,2) | 28 (20,7) | |
| Средний возраст (SD) | 61,5 (15,0) | 62,7 (15,3) | |
| IPI/NIVO Лечение N (%) | 15 (3,6) | 51 (38,8,8) | 44444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444. (%) (%) | 5 (1,1) | 29 (21,5) |
| Пол n (%) | |||
| Гнездовой | 175 (38,1) | 49 (36,3) | |
| Вилка | 284 (61,9) | 86 (63.7) | |
| Transcriptomics | RNA-seq | RNA-seq | |
| DNA mutation | WES | WES | |
Open in a separate window
2.
 1. Анализ последовательности WES и RNA-Seq
1. Анализ последовательности WES и RNA-Seq Данные WES были получены для опухолей и соответствующих нормальных образцов с глубиной охвата в среднем около 100×. Прочтения последовательностей были сопоставлены с эталонным геномом человека (hs37d5) с помощью прибора Burrows-Wheeler Aligner (BWA) [7], а перестройка вставок/делеций и повторная калибровка показателя качества были выполнены с помощью набора инструментов для анализа генома (GATK) [8]. Опухолеспецифические мутации были идентифицированы у Стрелки [9].] и MuTect [10] и были аннотированы для определения генетического контекста (т. е. несинонимичности, миссенс, сплайсинга) с использованием ANNOVAR [11]. Была включена дополнительная контекстуальная информация, в том числе частота аллелей в других исследованиях, таких как 1000 Genomes и проект NHLBI Exome Sequence Project, предсказания влияния функции in silico и наблюдаемые воздействия из баз данных, таких как ClinVar (http://www.ncbi.nlm.nih.gov). /clinvar/) (по состоянию на 1 марта 2016 г. ), Сборник соматических мутаций при раке (COSMIC) и Атлас генома рака (TCGA). Сигнатуры мутаций (изменения возможных последовательностей тринуклеотидов) подсчитывали и получали, как описано в Alexandrow et al. [12] как реализовано deconstructSigs [13]. Контроль качества WES включает в себя метрики считывания после каждого этапа анализа (дробные повторные считывания, фракционные сопоставленные считывания), оценку глубины покрытия в целевых регионах и общие сравнения генотипов между образцами для обеспечения надлежащего соответствия образцов. Мутации подсчитывались следующим образом: наблюдались в OR, специфичных для «Стрелки» (MuTect И «Стрелка-чувствительные»), предположительно изменяющие белок, и частота <1% на 1000 геномов.
), Сборник соматических мутаций при раке (COSMIC) и Атлас генома рака (TCGA). Сигнатуры мутаций (изменения возможных последовательностей тринуклеотидов) подсчитывали и получали, как описано в Alexandrow et al. [12] как реализовано deconstructSigs [13]. Контроль качества WES включает в себя метрики считывания после каждого этапа анализа (дробные повторные считывания, фракционные сопоставленные считывания), оценку глубины покрытия в целевых регионах и общие сравнения генотипов между образцами для обеспечения надлежащего соответствия образцов. Мутации подсчитывались следующим образом: наблюдались в OR, специфичных для «Стрелки» (MuTect И «Стрелка-чувствительные»), предположительно изменяющие белок, и частота <1% на 1000 геномов.
Данные секвенирования РНК также были получены для тех же образцов опухолей. Прочтения последовательностей были выровнены с эталонным геномом человека с учетом сплайсинга с использованием Tophat2 [14], что позволяет точно выравнивать последовательности по интронам. Выровненные последовательности были назначены экзонам с использованием пакета HTseq [15] для создания начальных подсчетов по регионам. Нормализация, моделирование экспрессии и тестирование различий выполнялись с использованием DESeq [16]. Контроль качества RNAseq включает в себя собственные скрипты и RSeqC [17] для проверки показателей количества прочтений, доли выравнивания, количества хромосомных выравниваний, показателей распределения экспрессии, а также анализа основных компонентов и иерархической кластеризации, чтобы гарантировать, что данные выборки представляют собой группу дизайна эксперимента. Данные полного экзома TCGA были загружены с веб-сайта NIH TCGA в марте 2016 г. [18]. Файл MAF был преобразован в VCF, а затем аннотирован, как описано для данных TCC. Мутации подсчитывали следующим образом: прогнозируется изменение белка и частота <1% на 1000 геномов. В этом исследовании использовался уровень 3 данных RNA-seq. Данные экспрессии RNA-seq были распределены между когортами TCGA и Moffitt с использованием функции Combat в пакете sva в R [19].
Выровненные последовательности были назначены экзонам с использованием пакета HTseq [15] для создания начальных подсчетов по регионам. Нормализация, моделирование экспрессии и тестирование различий выполнялись с использованием DESeq [16]. Контроль качества RNAseq включает в себя собственные скрипты и RSeqC [17] для проверки показателей количества прочтений, доли выравнивания, количества хромосомных выравниваний, показателей распределения экспрессии, а также анализа основных компонентов и иерархической кластеризации, чтобы гарантировать, что данные выборки представляют собой группу дизайна эксперимента. Данные полного экзома TCGA были загружены с веб-сайта NIH TCGA в марте 2016 г. [18]. Файл MAF был преобразован в VCF, а затем аннотирован, как описано для данных TCC. Мутации подсчитывали следующим образом: прогнозируется изменение белка и частота <1% на 1000 геномов. В этом исследовании использовался уровень 3 данных RNA-seq. Данные экспрессии RNA-seq были распределены между когортами TCGA и Moffitt с использованием функции Combat в пакете sva в R [19]. ].
].
2.2. Doublet Combination Therapy Candidates
Для прогнозирования лечения было включено в общей сложности 5894 лечения и их гены-мишени из базы данных SIGnatures DataBase DSigDB, [20], selleckchem.com (по состоянию на 1 марта 2016 г.), а также общеизвестные методы лечения иммунных контрольных точек [20]. ] в качестве кандидатов на терапию. Среди них 5845 препаратов были получены от DsigDB, 38 ингибиторов HDAC были получены от selleckchem.com, а 11 препаратов были известными препаратами для терапии иммунных контрольных точек.
Для начального скрининга для каждой терапии-кандидата был проведен анализ одиночной терапии, чтобы определить вероятные «исходные» терапии для формирования пула кандидатов на двойную комбинацию. Анализ отдельной терапии состоит из трех частей и был обобщен с использованием метода продукта Фишера (FPM). Для оценки потенциальной эффективности каждой отдельной терапии анализы мутаций, экспрессии и общей выживаемости пациентов (ОВ) использовались в качестве суррогата клинического результата. РНКсек и мутационный статус генов-мишеней терапии были первичными независимыми переменными в соответствующих моделях Cox PH с поправкой на возраст и 9Обработка 3165 IPI/NIVO и BRAF . Модели одиночной терапии для оценки связи между мутацией и ОС определяются как SMUTt=βMx+βaage+βBBRAF+βIIPI/NIVO, где x — показатель мутаций в генах-мишенях препарата-кандидата. BRAF является индикатором обработки ингибитором BRAF , а IPI/NIVO является индикатором любых ингибиторов контрольной точки. Для оценки связи между данными экспрессии генов и выживаемостью используется модель SPCt=βExPC+βaage+βBBRAF+βIIPI/NIVO, где xpC — первый главный компонент (PC1) данных экспрессии генов генов-мишеней лекарственного средства-кандидата. Поскольку лекарство часто нацелено на несколько генов, для обобщения и уменьшения размерности данных об экспрессии генов для генов, на которые нацелено лекарство, использовался анализ основных компонентов. PC1 объясняет максимальную степень дисперсии данных экспрессии для набора лекарств и использовался в анализе выживаемости и eQTL.
РНКсек и мутационный статус генов-мишеней терапии были первичными независимыми переменными в соответствующих моделях Cox PH с поправкой на возраст и 9Обработка 3165 IPI/NIVO и BRAF . Модели одиночной терапии для оценки связи между мутацией и ОС определяются как SMUTt=βMx+βaage+βBBRAF+βIIPI/NIVO, где x — показатель мутаций в генах-мишенях препарата-кандидата. BRAF является индикатором обработки ингибитором BRAF , а IPI/NIVO является индикатором любых ингибиторов контрольной точки. Для оценки связи между данными экспрессии генов и выживаемостью используется модель SPCt=βExPC+βaage+βBBRAF+βIIPI/NIVO, где xpC — первый главный компонент (PC1) данных экспрессии генов генов-мишеней лекарственного средства-кандидата. Поскольку лекарство часто нацелено на несколько генов, для обобщения и уменьшения размерности данных об экспрессии генов для генов, на которые нацелено лекарство, использовался анализ основных компонентов. PC1 объясняет максимальную степень дисперсии данных экспрессии для набора лекарств и использовался в анализе выживаемости и eQTL. Анализ eQTL проводили с использованием критерия суммы рангов Уилкоксона с использованием PC1 значений экспрессии целевых генов и индикаторов мутаций в целевых генах. При обнаружении мутации хотя бы в одном из целевых генов лекарственного средства (набора) для пациента статус мутации для этого пациента кодируется как 1 (в двоичном кодировании 0/1) для анализа eQTL. Если в генах-мишенях препарата (набора) мутации не обнаружено, то она кодируется как ноль. PC1 использовали в тесте суммы рангов Уилкоксона для оценки потенциальной связи между мутациями и экспрессией генов-мишеней в наборе лекарств. FPM был впервые использован для синтеза результатов трех анализов для каждого лекарства-кандидата в каждой когорте. Затем FPM использовался для создания сводки результатов двух когорт. p -значения, связанные со статистикой хи-квадрат из FPM, использовались для определения приоритета отдельного лечения. Частота ложных открытий (FDR) использовалась для корректировки множественных сравнений.
Анализ eQTL проводили с использованием критерия суммы рангов Уилкоксона с использованием PC1 значений экспрессии целевых генов и индикаторов мутаций в целевых генах. При обнаружении мутации хотя бы в одном из целевых генов лекарственного средства (набора) для пациента статус мутации для этого пациента кодируется как 1 (в двоичном кодировании 0/1) для анализа eQTL. Если в генах-мишенях препарата (набора) мутации не обнаружено, то она кодируется как ноль. PC1 использовали в тесте суммы рангов Уилкоксона для оценки потенциальной связи между мутациями и экспрессией генов-мишеней в наборе лекарств. FPM был впервые использован для синтеза результатов трех анализов для каждого лекарства-кандидата в каждой когорте. Затем FPM использовался для создания сводки результатов двух когорт. p -значения, связанные со статистикой хи-квадрат из FPM, использовались для определения приоритета отдельного лечения. Частота ложных открытий (FDR) использовалась для корректировки множественных сравнений. Обработки с FDR <0,05 были выбраны в качестве «исходных» терапий для сопряжения с каждым из оставшихся обработок для формирования пула кандидатов на дублет. Существует 37 терапий с FDR < 0,05: 26 одобренных FDA киназ, 7 иммунопрепаратов и 4 препарата HDAC. Мы также включили два клинически используемых препарата для лечения пациентов с меланомой: панобиностат и траметиниб, как часть 39обработка семян для создания дублетного пула. Сочетание каждой из обработок семян с каждой из оставшихся обработок из 5848 кандидатов дает в общей сложности 73 007 комбинаций в дублетном пуле.
Обработки с FDR <0,05 были выбраны в качестве «исходных» терапий для сопряжения с каждым из оставшихся обработок для формирования пула кандидатов на дублет. Существует 37 терапий с FDR < 0,05: 26 одобренных FDA киназ, 7 иммунопрепаратов и 4 препарата HDAC. Мы также включили два клинически используемых препарата для лечения пациентов с меланомой: панобиностат и траметиниб, как часть 39обработка семян для создания дублетного пула. Сочетание каждой из обработок семян с каждой из оставшихся обработок из 5848 кандидатов дает в общей сложности 73 007 комбинаций в дублетном пуле.
2.3. Модели перепрофилирования лекарств для дублетов
Для оценки потенциальных лечебных эффектов каждого кандидата на дублетную терапию была исследована связь между мутацией, экспрессией и общей выживаемостью пациентов, используемая в качестве суррогата клинического результата. RNAseq и мутационный статус генов-мишеней в рамках терапии были первичными независимыми переменными в соответствующих моделях Cox PH с поправкой на возраст и 9Обработка 3165 IPI/NIVO и обработка BRAF (уравнения (1) и (2)). Лечебное взаимодействие также было включено в модель. Анализ локусов количественных признаков экспрессии (eQTL) был выполнен для оценки потенциального влияния мутаций на транскрипцию с использованием теста Уилкоксона (уравнение (3)). Поскольку фактические механизмы лечебного действия через ДНК, РНК или их взаимодействие неясны, были сформулированы три модели/метода, основанные на различных предположениях. Метод 1 (уравнение (4)) объединяет все данные из (уравнений (1)–(3)). Метод 2 оценивает данные по экспрессии генов (уравнение (5)), а метод 3 оценивает наиболее значимые данные с минимумом p -значение среди 3 наборов доказательств (уравнения (1)–(3)) в каждой когорте (уравнение (6)). Подробности описаны ниже. Метод продукта Фишера использовался для объединения данных двух когорт. Дальнейшая фильтрация ( p <0,05) для каждой когорты была выполнена для Метода 2 и суммирована в Shiny App DRepMel. Анализы подмножеств были выполнены для пациентов с мутациями BRAF, NRAS или когорт Triple WT.
Лечебное взаимодействие также было включено в модель. Анализ локусов количественных признаков экспрессии (eQTL) был выполнен для оценки потенциального влияния мутаций на транскрипцию с использованием теста Уилкоксона (уравнение (3)). Поскольку фактические механизмы лечебного действия через ДНК, РНК или их взаимодействие неясны, были сформулированы три модели/метода, основанные на различных предположениях. Метод 1 (уравнение (4)) объединяет все данные из (уравнений (1)–(3)). Метод 2 оценивает данные по экспрессии генов (уравнение (5)), а метод 3 оценивает наиболее значимые данные с минимумом p -значение среди 3 наборов доказательств (уравнения (1)–(3)) в каждой когорте (уравнение (6)). Подробности описаны ниже. Метод продукта Фишера использовался для объединения данных двух когорт. Дальнейшая фильтрация ( p <0,05) для каждой когорты была выполнена для Метода 2 и суммирована в Shiny App DRepMel. Анализы подмножеств были выполнены для пациентов с мутациями BRAF, NRAS или когорт Triple WT.
Для оценки связи между общей выживаемостью пациентов и соматической мутацией (MUT), экспрессией генов с использованием PC1 и потенциальным влиянием мутации на гены-мишени были сформулированы следующие три уравнения. Для оценки выживаемости пациентов с мутацией в генах-мишенях каждого дублета:
SMUTt=βM1x1+βM2x2+βM3x1x2+βM4age+βM5BRAF+βM6IPI/NIVO
(1)
где x1 и x2 – индикаторы мутаций в генах-мишенях препаратов 1 и 2 соответственно. BRAF является индикатором лечения ингибитором BRAF, а IPI/NIVO является индикатором любых ингибиторов контрольных точек. Для оценки выживаемости пациентов с экспрессией в генах-мишенях каждого дуплета:
SPCt=βE1x1+βEE2x2+βE3x1x2+βM4age+βM5BRAF+βM6IPI/NIVO
(2)
где x1 и x2 – первый главный компонент данных экспрессии генов-мишеней препаратов 1 и 2 соответственно. Чтобы оценить потенциальное функциональное влияние мутаций на экспрессию генов, eQTL выполняется с помощью критерия суммы рангов Уилкоксона. PC1 значений экспрессии генов-мишеней использовали с индикаторами мутаций в генах-мишенях обоих препаратов.
PC1 значений экспрессии генов-мишеней использовали с индикаторами мутаций в генах-мишенях обоих препаратов.
eQTL:PC11~ x1 целевых генов в лекарстве 1 PC12~ x2 целевых генов в лекарстве
(3)
Для повышения надежности вывода был проведен анализ с использованием двух независимых когорт пациентов с меланомой: когорты TCGA (N = 459) и когорты с меланомой Моффита (N = 135). Метод продукта Фишера использовался для синтеза результатов каждого из приведенных выше анализов (уравнения (1)–(3)) со следующими обозначениями: p(β) представляет собой p -значение коэффициента β в уравнениях 1 или 2. или p — значение анализа eQTL (3).
M = модель «Мутация», E = модель экспрессии.
«1» = препарат 1, «2» = препарат 2
«t» = когорта TCGA, «m» = когорта Моффита
модели были сформулированы на основе различных предположений.
Метод 1 объединяет все данные из уравнений (1)–(3):
Meta P (включает все восемь терминов из обеих групп)
χ2k 2~−2∑(lnPβM1t+lnPβM2t+lnPβM3t+lnPβE1t+lnPβE2t+lnPβE3t+lnPeQTL1t+lnPeQTL2t+lnPβM1m+lnPβM2m+lnPβM3m+lnPβE1m+lnPβE2m+lnPβE3m+lnPeQTL1m+lnPeQTL2m)
(4)
Метод 2 оценивает свидетельство ассоциации, используя только данные экспрессии: CombinedPEExpression (включает основные эффекты из уравнения выражения (2)).
χ2k 2~−2∑lnPβE1t+lnPβE2t+lnPβE1m+lnPβE2m
(5)
Поскольку механизм действия может различаться в зависимости от лечения, поэтому в каждой когорте минимальное значение p теста среди 3 тестов (Уравнения (1)–(3)).
Метод 3 объединяет минимальное значение p тестов между двумя когортами. Например, если мин. p -значение взято из той же модели, скажем, модели SMUTt в когортах TCGA и Moffitt, тогда используется термин взаимодействия p -значения.
χ2k2~−2∑lnPβM3t+lnPβM3m
(6)
2.4. Потенциальная мишень TME с помощью предсказанных двойных терапий
Потенциальная мишень TME с помощью предсказанных двойных терапий была сделана с использованием данных scRNA-seq 28 078 одиночных клеток из 43 образцов пациентов [5]. Все анализы и приложение Shiny были выполнены и реализованы с использованием R.
Предсказанные дублеты по Методу 2 версии 1.0 Drepmel доступны для визуализации с помощью приложения Shiny на http://drepmel. moffitt.org/, которое будет поддерживаться не менее 3 лет (обращайтесь по адресу gro.ttiffom@nospmoht .yrahcaz или [email protected] по техническим вопросам). Код R, определяющий серверную логику приложения Shiny, доступен в дополнительном файле S1. Код R, управляющий макетом и внешним видом приложения, доступен в дополнительном файле S2.
moffitt.org/, которое будет поддерживаться не менее 3 лет (обращайтесь по адресу gro.ttiffom@nospmoht .yrahcaz или [email protected] по техническим вопросам). Код R, определяющий серверную логику приложения Shiny, доступен в дополнительном файле S1. Код R, управляющий макетом и внешним видом приложения, доступен в дополнительном файле S2.
Ввод данных для приложения включает в себя два раскрывающихся меню лечения и переключатели для выбора (под)группы пациентов, соответствующих основным генотипам меланомы (All, NRAS, BRAF, Triple WT). Два дополнительных раскрывающихся меню предоставляют целевые гены для выбора в каждой обработке для тепловых карт экспрессии одного гена, чтобы понять потенциальный эффект обработки в TME.
Приложение Shiny включает вкладку для введения, метод и следующие вкладки результатов:
В таблицах верхних комбинаций дублетов обобщены общие результаты и результаты для каждой из основных групп генотипов меланомы.
TME: тепловая карта и график скрипки Выделите потенциальные целевые популяции клеток для каждой терапии
На вкладке мутации и выживаемости отображаются графики Каплана-Мейера общей выживаемости, основанные на статусе мутаций в генах-мишенях выбранных дублетов в каждой когорте .
На вкладке PC1 и выживаемость показаны таблицы генов и загрузок PC1 в наборах целевых генов каждого лечения для каждой когорты, а также графики КМ для PC1 и общей выживаемости для каждого лечения в обеих когортах. Значения PC1 дихотомированы по медиане.
На вкладке eQTL отображаются диаграммы экспрессии генов в обеих когортах по статусу мутации в целевых генах. Он также отображает сводную статистику депакетного выражения в логарифмической шкале.

Результаты методов 1, 2 и 3 включены в дополнительные файлы S3–S5. Верхние комбинации включают вероятных кандидатов. Для общего анализа лучшие комбинации включают известные эффективные методы лечения (анти-PD1), вероятные (Lag3, нилотиниб) и дополнительные комбинации методов лечения, которые могут быть дополнительно исследованы (дополнительный файл S4). Надежные результаты между когортами TCGA и Moffitt для пациентов с ограниченными вариантами лечения (пациенты с NRAS или тройным WT) дают краткий список кандидатов для дальнейшего исследования. 52 предсказанные комбинации для подгруппы NRAS содержат несколько интересных кандидатов. Лучшие кандидаты, сочетающие LAG3 и клиохинол, показывают согласованные результаты между двумя когортами пациентов (1). Эта комбинация, хотя и неожиданная, может предложить новые возможности для лечения меланомы. Клиохинол оказывает влияние на протеасомы, а также на метаболизм меди и цинка и может изменять транскрипционную активность как в раковых, так и в иммунных клетках [21,22]. Вполне возможно, что эти широко направленные эффекты на состояние транскрипции опухоли могут повысить чувствительность к более широко используемым иммунотерапевтическим средствам, таким как антитело против LAG3. Наша группа уже продемонстрировала, что иммунотерапия может использоваться последовательно с таргетной терапией для обеспечения долгосрочного противоопухолевого эффекта на моделях меланомы у мышей. В данном случае эти эффекты обусловлены как модуляцией передачи сигналов в опухоли, так и перепрограммированием иммунного микроокружения [23].
52 предсказанные комбинации для подгруппы NRAS содержат несколько интересных кандидатов. Лучшие кандидаты, сочетающие LAG3 и клиохинол, показывают согласованные результаты между двумя когортами пациентов (1). Эта комбинация, хотя и неожиданная, может предложить новые возможности для лечения меланомы. Клиохинол оказывает влияние на протеасомы, а также на метаболизм меди и цинка и может изменять транскрипционную активность как в раковых, так и в иммунных клетках [21,22]. Вполне возможно, что эти широко направленные эффекты на состояние транскрипции опухоли могут повысить чувствительность к более широко используемым иммунотерапевтическим средствам, таким как антитело против LAG3. Наша группа уже продемонстрировала, что иммунотерапия может использоваться последовательно с таргетной терапией для обеспечения долгосрочного противоопухолевого эффекта на моделях меланомы у мышей. В данном случае эти эффекты обусловлены как модуляцией передачи сигналов в опухоли, так и перепрограммированием иммунного микроокружения [23]. Тщательная доклиническая оценка комбинаций лекарственных средств, выбранных из таких инструментов, как DRepMel, может привести к созданию надежного набора перепрофилированных комбинаций лекарственных средств для будущей клинической оценки. Результаты ТМЭ показывают, что каждая терапия, вероятно, нацелена на разные клеточные популяции: лимфоидные и миелоидные, соответственно. Это дает представление о том, как могут работать возможные комбинации. Надежные прогнозы также предоставляются для подгруппы пациентов с меланомой по шкале BRAF. Инструмент DRepMel предоставляет полезный вычислительный ресурс с надежными выводами для генерации гипотез. Это также дает представление о потенциальном воздействии лечения на TME для дальнейшего изучения.
Тщательная доклиническая оценка комбинаций лекарственных средств, выбранных из таких инструментов, как DRepMel, может привести к созданию надежного набора перепрофилированных комбинаций лекарственных средств для будущей клинической оценки. Результаты ТМЭ показывают, что каждая терапия, вероятно, нацелена на разные клеточные популяции: лимфоидные и миелоидные, соответственно. Это дает представление о том, как могут работать возможные комбинации. Надежные прогнозы также предоставляются для подгруппы пациентов с меланомой по шкале BRAF. Инструмент DRepMel предоставляет полезный вычислительный ресурс с надежными выводами для генерации гипотез. Это также дает представление о потенциальном воздействии лечения на TME для дальнейшего изучения.
Открыть в отдельном окне
DRepMel прогнозирует, что комбинированное лечение LAG3 и клиохинола (HL60 ниже) может быть рассмотрено для пациентов с меланомой с мутациями NRAS. Сигналы устойчивы и устойчивы в когортах ( A , B ) TCGA и ( C , D ) Moffitt. ( E , F ) Каждая терапия, вероятно, нацелена на разные клеточные популяции, т.е. T/NK и миелоидные, как показано с использованием данных scRNA-seq.
( E , F ) Каждая терапия, вероятно, нацелена на разные клеточные популяции, т.е. T/NK и миелоидные, как показано с использованием данных scRNA-seq.
Открыть в отдельном окне
DRepMel прогнозирует, что комбинированное лечение LAG3 и азатиоприна (снижение MCF7) может быть рассмотрено для пациентов с меланомой с мутациями BRAF. Сигналы устойчивы и устойчивы в когортах ( A , B ) TCGA и ( C , D ) Moffitt. ( E , F ) Каждая терапия, вероятно, нацелена на разные отделы TME.
Благодарим Родриго Карвахаля и Гильермо Гонсалес-Кальдерон за техническую поддержку.
Следующую вспомогательную информацию можно загрузить по адресу: https://www.mdpi.com/article/10.3390/cells11182894/s1, дополнительный файл S1 — файл Server.R: код R, содержащий инструкции по созданию приложения. Определяет серверную логику приложения Shiny. Включает создание функций, которые отображают пользовательский ввод в различные виды вывода; Дополнительный файл S2 — файл ui. R: код R, управляющий макетом и внешним видом приложения. Функции R — это оболочки для HTML, CSS и JS для разработки адаптивных приложений; Дополнительный файл S3 — результаты метода 1: файл Excel с 3 рабочими листами: результаты, словарь данных, сокращения; Дополнительный файл S4 — результаты метода 2: файл Excel с 3 рабочими листами: результаты, словарь данных, сокращения; Дополнительный файл S5 — результаты метода 3: файл Excel с 3 рабочими листами: результаты, словарь данных, сокращения.
Щелкните здесь для получения файла с дополнительными данными. (2.9M, zip)
Эта работа была поддержана наградой Moffitt SKIN SPORE (P50 CA168536) DRP, премией DRP Core Innovation Project, а также частично поддержана Национальным институтом рака через грант поддержки онкологического центра Moffitt (P30 -CA076292).
Ю.А.К. придумал концепцию и разработал DRepMel. З.Дж.Т. разработал приложение Shiny и закодировал предсказание комбинации наркотиков. Дж.К.Т. выполнили анализы WES. JL закодировал визуализацию TME. З.К. извлекли данные DsigDB для прогнозирования лечения. Ю.З. выполнили анализ РНК-секвенирования когортного набора данных Моффитта. Э.А.В. выполнили QC и разделили данные секвенирования РНК между когортами Moffitt и TCGA. Н.А. и З.Е. абстрагированная медицинская информация для когорты Моффита. Y.A.C., J.K.T. и З.Дж.Т. составил рукопись. К.С.М.С. и А.С.Т. пересмотрела проект и предоставила обратную связь. Все авторы прочитали и согласились с опубликованной версией рукописи.
Это исследование (MCC# 19147) проводилось в Онкологическом центре Моффитта в соответствии с признанными этическими принципами (например, Хельсинкская декларация, CIOMS, Белмонтский отчет, Общее правило США) и было одобрено Чесапикским институциональным наблюдательным советом (IRB). Отказ от согласия был предоставлен IRB (IRB # 00000790). IRB определил, что наше исследование соответствует критериям отказа от согласия в соответствии с 45 CFR 46.116(d) и отказа от авторизации HIPAA в соответствии с 45 CFR 164. 512(i)(2). IRB предоставил отказ от согласия и отказ от авторизации HIPAA.
Первоначально пациенты дали согласие на использование протокола Total Cancer Care Protocol, институционального биорепозитория Moffitt Cancer Center (MCC#14690; Advarra IRB Pro00014441). Работа по исследованию проводилась в соответствии с протоколом MCC# 19147. Chesapeake IRB определил, что наше исследование соответствует критериям отказа от согласия в соответствии с 45 CFR 46.116(d) и отказа от авторизации HIPAA в соответствии с 45 CFR 164.512(i)(2) . IRB предоставил отказ от согласия и отказ от авторизации HIPAA.
Данные и результаты, полученные в ходе этого исследования, доступны в виде дополнительных файлов и доступны по запросу через веб-сайт. Все коды R загружаются в виде дополнительных файлов server.R и ui.R. Руководство пользователя также доступно на веб-сайте.
Авторы заявляют об отсутствии конфликта интересов.
Примечание издателя: MDPI сохраняет нейтралитет в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
1. Цао Х., Чин Л., Гаррауэй Л.А., Фишер Д.Е. Меланома: от мутаций к медицине. Гены Дев. 2012; 26:1131–1155. doi: 10.1101/gad.1
2. Li P., Huang C., Fu Y., Wang J., Wu Z., Ru J., Zheng C., Guo Z. , Chen X., Zhou W. и соавт. Крупномасштабное исследование и анализ комбинаций лекарств. Биоинформатика. 2015;31:2007–2016. дои: 10.1093/биоинформатика/btv080. [PubMed] [CrossRef] [Google Scholar]
3. Wildenhain J., Spitzer M., Dolma S., Jarvik N., White R., Roy M., Griffiths E., Bellows D.S., Wright G.D., Tyers M. , Прогнозирование синергизма химико-генетических взаимодействий с помощью машинного обучения. Сотовая система 2015;1:383–395. doi: 10.1016/j.cels.2015.12.003. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
4. Preuer K., Lewis R.P.I., Hochreiter S., Bender A., Bulusu K.C., Klambauer G. DeepSynergy: прогнозирование синергии противораковых препаратов с Deep Обучение. Биоинформатика. 2018;34:1538–1546. дои: 10.1093/биоинформатика/btx806. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
5. Smalley I., Chen Z., Phadke M., Li J., Yu X., Wyatt C., Evernden B., Messina J.L., Сарнаик А., Сондак В.К. и др. Одноклеточная характеристика иммунного микроокружения меланомы головного мозга и лептоменингеальных метастазов. клин. Рак рез. 2021;27:4109–4125. doi: 10.1158/1078-0432.CCR-21-1694. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
6. Wang X., Yu X., Krauthammer M., Hugo W., Duan C., Kanetsky P.A., Teer J.K., Thompson Z.J., Kalos D. ., Цай К.Ю. и соавт. Связь мутации MUC16 с бременем мутаций опухоли и ее прогностическим значением при меланоме кожи. Эпидемиол рака. Биомарк. Пред. 2020;29: 1792–1799. doi: 10.1158/1055-9965.EPI-20-0307. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
7. Ли Х., Дурбин Р. Быстрое и точное выравнивание коротких чтений с помощью преобразования Берроуза-Уилера. Биоинформатика. 2009; 25:1754–1760. doi: 10.1093/биоинформатика/btp324. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
8. ДеПристо М.А., Бэнкс Э., Поплин Р., Гаримелла К.В., Магуайр Дж.Р., Хартл К., Филиппакис А.А., дель Анхель Г., Ривас М.А. , Ханна М. и др. Основа для обнаружения вариаций и генотипирования с использованием данных секвенирования ДНК следующего поколения. Нац. Жене. 2011;43:491–498. doi: 10.1038/ng.806. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Рудницкий С., Захави И. Антибиотикопрофилактика для предотвращения инфекционного эндокардита. Харефуа. 1990; 119: 275–277. [PubMed] [Google Scholar]
10. Цибульскис К., Лоуренс М.С., Картер С.Л., Сиваченко А., Джаффе Д., Сугнез С., Габриэль С., Мейерсон М., Ландер Э.С., Гетц Г. Чувствительное обнаружение соматические точечные мутации в неочищенных и гетерогенных образцах рака. Нац. Биотехнолог. 2013; 31: 213–219. doi: 10.1038/nbt.2514. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. Ван К. , Ли М., Хаконарсон Х. ANNOVAR: Функциональная аннотация генетических вариантов на основе данных высокопроизводительного секвенирования. Нуклеиновые Кислоты Res. 2010;38:e164. doi: 10.1093/nar/gkq603. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
12. Александров Л.Б., Ник-Зайнал С., Ведж Д.К., Апарисио С.А., Бехжати С., Бьянкин А.В., Бигнелл Г.Р., Болли Н., Борг А. ., Borresen-Dale A.L., et al. Признаки мутационных процессов при раке человека. Природа. 2013; 500:415–421. doi: 10.1038/nature12477. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Маршалл А.Х., Матилла А., Поллак Д.Дж. Критика и тематическое исследование узловой склерозирующей болезни Ходжкина. Дж. Клин. Патол. 1976; 29: 923–930. doi: 10.1136/jcp.29.10.923. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
14. Trapnell C., Pachter L., Salzberg S.L. TopHat: обнаружение соединений сплайсинга с помощью RNA-Seq. Биоинформатика. 2009;25:1105–1111. doi: 10.1093/биоинформатика/btp120. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
15. Андерс С., Пил П.Т., Хубер В. HTSeq — платформа Python для работы с высокопроизводительными данными секвенирования. Биоинформатика. 2015; 31: 166–169. doi: 10.1093/биоинформатика/btu638. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Андерс С., Хубер В. Анализ дифференциальной экспрессии для данных подсчета последовательностей. Геном биол. 2010;11:R106. doi: 10.1186/gb-2010-11-10-r106. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
17. Ван Л., Ван С., Ли В. RSeQC: Контроль качества экспериментов по секвенированию РНК. Биоинформатика. 2012;28:2184–2185. doi: 10.1093/биоинформатика/bts356. [PubMed] [CrossRef] [Академия Google]
18. Набор данных TCGA SKIN DNAseq. [(по состоянию на 1 марта 2016 г.)]; Доступно в Интернете: https://tcga-dta.nci.nih.gov/tcgafiles/ftp_auth/distro_ftpusers/anonymous/tumor/skcm/gsc/broad.mit.edu/illuminaga_dnaseq_automated/muttions/broad. mit.edu_SKCM.IlluminaGA_DNASeq_automated.Level_2 .1.5.0/SKCM_pairs.aggregated.capture.tcga.uuid.automated.somatic.maf
19. Leek J.T., Johnson W.E., Parker H.S., Jaffe A.E., Storey J.D. Пакет sva для удаления пакетных эффектов и других нежелательных изменений в высокопроизводительные эксперименты. Биоинформатика. 2012; 28:882–883. дои: 10.1093/биоинформатика/bts034. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
20. Ю М., Шин Дж., Ким Дж., Райалл К.А., Ли К., Ли С., Чон М., Кан Дж., Tan AC DSigDB: база данных сигнатур лекарств для анализа набора генов. Биоинформатика. 2015;31:3069–3071. doi: 10.1093/биоинформатика/btv313. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
21. Хан Р., Хан Х., Абдулла Ю., Доу К.П. Возможность повторного использования клиохинола для лечения рака. Недавний Пэт. Противораковый препарат Дисков. 2020;15:14–31. дои: 10.2174/157486662002270
22. Treiber C., Simons A. , Strauss M., Hafner M., Cappai R., Bayer T.A., Multhaup G. Клиохинол опосредует поглощение меди и противодействует ее оттоку. Активность белка-предшественника амилоида при болезни Альцгеймера. Дж. Биол. хим. 2004; 279:51958–51964. doi: 10.1074/jbc.M407410200. [PubMed] [CrossRef] [Google Scholar]
23. Phadke M.S., Chen Z., Li J., Mohamed E., Davies M.A., Smalley I., Duckett D.R., Palve V., Czerniecki B.J., Forsyth P.A., et al. др. Таргетная терапия, назначаемая после анти-PD-1, приводит к пролонгированным ответам на моделях мышиной меланомы благодаря устойчивому противоопухолевому иммунитету. Рак Иммунол. Рез. 2021;9: 554–567. doi: 10.1158/2326-6066.CIR-20-0905. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
PLOS Medicine
6 сентября 2022 г.
Редакторы PLOS Medicine обсуждают значение парашютной науки и стратегии для решения этой проблемы во всем мире.
Изображение предоставлено: Gerd Altmann, Pixabay
15.09.2022
Исследовательская статья
В исследовании случай-контроль Jiangwei Sun и его коллеги изучают взаимосвязь между стационарными инфекциями и риском диагностики болезни Альцгеймера. , болезнь Паркинсона и БАС с использованием данных общенационального регистра от 1970-2016 в Швеции.
Изображение предоставлено Стивеном ХВГ, Unsplash
.Недавно опубликованные статьи
- Связь травм после начала приема опиоидов по рецепту с риском побочных эффектов, связанных с опиоидами, среди пожилых получателей Medicare в США: вложенное исследование случай-контроль
- Траектории индекса массы тела у детей и связь с хронической болезнью почек у взрослых в Дании: популяционное когортное исследование
- Воздействие PM 9, связанного с лесными пожарами3418 2,5 и местная смертность от рака в Бразилии с 2010 по 2016 год: ретроспективное исследование
Просмотреть все статьи
Обзор выпусков за 2004–2022 гг.
15.09.2022
Перспектива .
Изображение предоставлено Кеном Кахири, Unsplash
.15.09.2022
Исследовательская статья
В ходе когортного исследования Эндрю Эдмондс и его коллеги изучают клинические и программные результаты воздействия ВИЧ-инфицированных детей на лечение в географически разных клиниках с 1997 по 2021 год.
Изображение предоставлено: CDC, CDC
06.09.2022
Исследовательская статья
Цянь Ян и его коллеги исследуют связь между бессонницей и мертворождением, невынашиванием беременности, гестационным диабетом, гипертоническими расстройствами беременности, перинатальной депрессией, преждевременными родами, а также низкой и высокой массой тела при рождении.
Изображение предоставлено: Джоуи Томпсон, Unsplash
08.09.2022
Исследовательская статья
Мариам О. Фофана и ее коллеги исследуют распространенность серотипа SARS-CoV-2 в городских трущобах Бразилии.
Изображение предоставлено Фернандо Сантосом, Unsplash
.08.09.2022
Исследовательская статья
Кэролайн А. Кроутер и ее коллеги исследуют, снижают ли более или менее жесткие целевые гликемические показатели для женщин с гестационным сахарным диабетом материнскую и перинатальную заболеваемость в Новой Зеландии.
Изображение предоставлено: Москва, Pixabay
01. 09.2022
Научная статья
Нона Саргисян и ее коллеги исследуют риск детского рака у детей, родившихся после переноса замороженных-размороженных эмбрионов в Дании, Финляндии, Норвегии и Швеции.
Изображение предоставлено: Елена Έλενα Kontogianni Κοντογιάννη, Pixabay
01.09.2022
Исследовательская статья
Mie Agermose Gram et al оценивают эффективность вакцины против COVID-19инфекционное заболевание.
Изображение предоставлено: Муфид Меджнун, Unsplash
01.09.2022
Исследовательская статья
Leen Farouki et al. обсудить глобальную распространенность и распространение калечащих операций на женских половых органах.
Изображение предоставлено: Крисси Кремер, Unsplash
02.09.2022
Исследовательская статья
Клара Понс-Дюран и ее коллеги изучают клиническую малярию во время беременности.
Изображение предоставлено: Мика Мами, Pixabay
Сложные медицинские ассоциации в различных контекстах требуют индивидуальных и основанных на правах человека решений
В редакционной статье приглашенные редакторы Пол Шпигель, Терри Макговерн и Кол Викрэмидж обсуждают специальный выпуск.
Страдания и потребность в уходе беженцев-рохинджа и лиц, обеспечивающих уход, в Бангладеш
Меган Доэрти и ее коллеги сообщают о болезнях, страданиях и потребностях в лечении среди беженцев-рохинджа и лиц, обеспечивающих уход, в Бангладеш.
Медицинские факторы, связанные с риском мертворождения у мигрантов в Западной Австралии
Марьям Мозуни и ее коллеги выявили более высокий риск интранатального мертворождения у женщин-мигрантов в Австралии по сравнению с женщинами, родившимися в Австралии.
Изучение источников незащищенности эфиопских женщин оромо и сомалийцев, родивших в лагере беженцев Какума
Эмбер Лалла и колл. изучить источники незащищенности эфиопских и сомалийских женщин, родивших в лагере беженцев Какума.
Сборник по гармонизации регулирования
В этом специальном сборнике представлен обзор Инициативы по гармонизации регулирования лекарственных средств в Восточной Африке, описывается ее прогресс и раскрывается дорожная карта инициативы на следующий этап.
Употребление психоактивных веществ, злоупотребление ими и зависимость
В этом специальном выпуске рассказывается о различных сообществах, которые работают для поддержки людей и групп населения, пострадавших от употребления психоактивных веществ и злоупотребления ими.
Здоровье и питание матери и ребенка
В этом специальном выпуске за 2019 г. подчеркивается важность питания для здоровья матери и ребенка.
Петля отрицательной обратной связи miR-3648/FRAT1-FRAT2/c-Myc модулирует метастазирование и инвазию клеток рака желудка
Петля отрицательной обратной связи miR-3648/FRAT1-FRAT2/c-Myc модулирует метастазирование и инвазию рака желудка клетки
Скачать PDF
Скачать PDF
- Артикул
- Открытый доступ
- Опубликовано:
- Вэймэй Тан 1 ,
- Мяомяо Пей 1 ,
- Цзяин Ли 1 ,
- Nanzhu Xu 1 ,
- Wushuang Xiao 1 ,
- Zhen Yu 1 ,
- Jieming Zhang 1 ,
- Linjie Hong 1 ,
- Zheng Guo 2 ,
- Jianjiao Lin 3 ,
- Weiyu Dai 1 ,
- Yizhi Xiao 1 ,
- Xiaosheng Wu 1 ,
- Guangnan Liu 1 ,
- Fachao Zhi ORCID: orcid.org/0000-0001-8674-4737 1,3 ,
- Гуоксин Ли Orcid: orcid.org/0000-0003-2773-7048 4 ,
- Jing Xiong 1 ,
- Ye Chen 5 ,
- HUI ,
- HUI ZHEN ,
- HUI ZHEN ,
- HUI ,
- HUI ,
- HUI . ORCID: orcid.org/0000-0002-7164-0588 3 ,
- Аймин Ли ORCID: orcid.org/0000-0002-0797-8496 1 ,
- Сайд Лю ORCID: orcid.org/0000-0002-7138-0245 1,3 и
- …
- Цзиде Ван ORCID: orcid.org/0000-0002-0988-9454 1,3
Онкоген (2022)Цитировать эту статью
Сведения о показателях
Субъекты
- Рак желудка
- Онкогены
Abstract
Хотя аномальная экспрессия miRNAs в раковых клетках является общепризнанным явлением, молекулярные механизмы, лежащие в основе прогрессирования miR-3648 и метастазирования при раке желудка (GC), остаются неясными. Экспрессия миР-3648 подавляется, и ее эктопическая экспрессия в клетках GC значительно подавляет клеточную пролиферацию и метастазирование. Механистический анализ показал, что miR-3648 непосредственно нацеливается на FRAT1 или FRAT2 и ингибирует опосредованную FRAT1 или FRAT2 инвазию и подвижность in vitro и in vivo. Более того, FRAT1 физически взаимодействует с FRAT2. Кроме того, сверхэкспрессия FRAT1 способствует инвазии клеток GC, тогда как siRNA-опосредованная репрессия FRAT2 в клетках GC со сверхэкспрессией FRAT1 обращает вспять его инвазивный потенциал. Кроме того, миР-3648 инактивировала сигнальный путь Wnt/β-катенин, подавляя FRAT1 и FRAT2 при GC. Интересно, что c-Myc, нижестоящий эффектор передачи сигналов Wnt/β-catenin, также подавлялся сверхэкспрессией miR-3648. В свою очередь, c-Myc негативно регулирует экспрессию miR-3648, связываясь с промотором miR-3648. Кроме того, уровни экспрессии миР-3648 отрицательно коррелировали с экспрессией c-Myc, FRAT1 и FRAT2 в свежих образцах желудка. Наши исследования показывают, что miR-3648 действует как опухолесупрессирующая miRNA и что петля отрицательной обратной связи miR-3648/FRAT1-FRAT2/c-Myc может быть критическим регулятором прогрессирования GC.
История вопроса
Рак желудка (РЖ) является пятым по распространенности видом рака и третьей по частоте причиной смерти от рака во всем мире [1]. Несмотря на недавний прогресс в выявлении и лечении раннего РЖ, долгосрочная выживаемость пациентов с распространенным РЖ низкая [2]. Таким образом, срочно необходимы идентификация диагностических биомаркеров и лучшее понимание молекулярных механизмов, лежащих в основе прогрессирования РЖ.
Считается, что микроРНК (миРНК или миР) длиной примерно 22 нуклеотида регулируют экспрессию генов путем связывания с 3′-нетранслируемой областью (3′-UTR) мРНК, что приводит к деградации мРНК или блокированию трансляции мРНК [ 3, 4]. Сообщалось, что различные микроРНК участвуют в прогрессировании GC. Например, эктопическая экспрессия miR-647 значительно ингибировала мРНК и белок SRF, а экспрессия miR-647 отрицательно ассоциировалась с мРНК SRF [3]. Наши предыдущие отчеты подтвердили, что miR-646 является супрессором опухоли для GC, который непосредственно регулирует экспрессию FOXK1 [4].
миР-3648 является менее изученной миРНК. Он расположен на хромосоме 21p11.2 [5] и участвует в нескольких типах рака. Сообщения показали, что miR-3648 модулирует пролиферацию клеток рака предстательной железы путем нацеливания на APC2 [6]. Кроме того, сверхэкспрессия миР-3648 регулирует миграцию и инвазию клеток рака мочевого пузыря [7]. Однако биологическая функция и основные механизмы miR-3648 при РЖ остаются неясными.
Сигнальный путь Wnt/β-catenin является одним из наиболее важных внутриклеточных путей, контролирующих прогрессирование рака [8]. Передача сигналов Wnt/β-катенина передается через семейство рецепторов Frizzled и корецептор LRP5/LRP6 в сигнальный каскад β-катенина [9]. , 10]. PAR-1, CKI epsilon, FRAT1 и FRAT2 являются положительными регуляторами канонического пути Wnt, тогда как APC, AXIN1, AXIN2 и PPARg являются отрицательными регуляторами [9, 11]. Ядерный комплекс, состоящий из TCF/LEF [12], активирует транскрипцию канонических генов-мишеней Wnt, таких как c-Myc [13], JUN [14, 15], циклин D1 [16, 17] и HIF-1a. [18]. Некоторые сообщения показывают, что miRNAs регулируют гены-мишени в сигнальном пути Wnt/β-catenin. Например, miRNA-194 активирует сигнальный путь Wnt/β-catenin в GC путем нацеливания на SUFU [19].]. miR-504 подавляет мезенхимальный фенотип опухолей путем прямого нацеливания на FZD7-опосредованный путь Wnt-β-catenin [20]. Однако, не совсем понятно, участвует ли miR-3648 в пути передачи сигналов Wnt при GC.
Здесь мы представляем доказательства того, что миР-3648 является потенциальным супрессором опухоли, который непосредственно нацеливается на FRAT1-FRAT2 (часто реаранжируется при прогрессирующих Т-клеточных лимфомах-1 и 2) для инактивации пути Wnt/β-катенин. Кроме того, мы наблюдали, что miR-3648 ингибировалась онкогенным транскрипционным фактором, c-Myc-a Wnt/β-catenin-downstream геном-мишенью, тем самым образуя петлю отрицательной обратной связи между miR-3648/FRAT1-FRAT2/c-Myc, тем самым усиливая прогрессирование GC.
Результаты
Понижение экспрессии миР-3648 связано с плохим прогнозом при РЖ
Сначала мы провели полногеномный скрининг для выявления микроРНК, связанных с клиническими исходами у пациентов с РЖ, в соответствии со строгими статистическими критериями ( P < 0,01) в базе данных TCGA с использованием онлайн-ресурсов oncomir (http://www.oncomir.org/). Мы обнаружили, что 26 микроРНК были связаны с общей выживаемостью (ОВ).
Высокая экспрессия 15 микроРНК предсказывает плохой прогноз и действует как потенциальный онкоген у пациентов с РЖ (дополнительная таблица 2), тогда как низкая экспрессия 11 микроРНК предсказывает неблагоприятный прогноз и действует как предполагаемый супрессор опухоли (дополнительная таблица 3). Среди них экспрессия миР-3648 была связана как с краткосрочной, так и с долгосрочной ОВ пациентов (рис. 1А). Более того, экспрессия миР-3648 была связана с кратковременной безрецидивной выживаемостью пациентов, но экспрессия миР-3648 не была связана с безрецидивной выживаемостью с течением времени (рис. 1Б).
A , B Анализ общей выживаемости (ОВ) и безрецидивной выживаемости (БРВ) проводили на основе кривых Каплана-Мейера в базе данных онкомир. C Экспрессию миР-3648 определяли с помощью количественной ПЦР в 82 тканях желудка человека, которая была нормализована по отношению к эндогенному контролю РНК U6. NT нормальные ткани слизистой оболочки, GC GC ткани. *** P < 0,01, пара т -тест. D Относительные уровни miR-3648 в GC и нормальных тканях, измеренные с помощью количественной ПЦР, были показаны с использованием графика водопада. Столбики погрешностей представляют собой среднее ± SD из трех независимых экспериментов. E Экспрессия миР-3648 в восьми линиях эпителиальных клеток желудка по данным количественной ПЦР. Односторонний ANOVA и критерий множественного сравнения Dunnett T3. **** P < 0,001 между нормальными и раковыми эпителиальными клетками желудка. Результат представлен как среднее ± SD трех независимых экспериментов. F , G Взаимосвязь между экспрессией миР-3648 и клиническими стадиями или локальной инвазией. *** P < 0,01; **** P < 0,001. Значения P оценивали с помощью двустороннего непарного t -критерия Стьюдента. H ISH-анализ уровня экспрессии миР-3648 в нормальной слизистой оболочке желудка и тканях GC. Шкала баров, 100 мкм в H .
Изображение полного размера
Во-вторых, мы сосредоточились на низкой экспрессии 11 микроРНК: miR-1306-3p, miR-3648, miR-2115-3p, let-7f-5p [21], miR-7-5p [ 22], миР-96-3p, миР-942-5p [23], миР-135b-3p, миР-423-5p [24], миР-221-3p [25], миР-15b-5p [26]. Используя базу данных PubMed-MEDLINE, мы обнаружили, что miR-1306-3p, miR-2115-3p, miR-96-3p, miR-135b-3p и miR-3648 еще не были зарегистрированы в GC. Мы обнаружили экспрессию этих пяти микроРНК в 30 тканях желудка человека с помощью количественной ПЦР. За исключением повышенной экспрессии miR-135-3p, экспрессия остальных четырех микроРНК была снижена в тканях GC (T), чем в нормальной слизистой оболочке желудка (N) (дополнительная рис. 1). Среди них miR-3648 подвергалась наиболее глубокому подавлению. Таким образом, мы задались вопросом, может ли дерегуляция миР-3648 участвовать в развитии и прогрессировании опухоли. Затем мы измерили экспрессию miR-3648 в 82 образцах желудка и подтвердили результаты вышеупомянутых исследований (9).3697 P = 0,002, рис. 1В, Г).
В-третьих, мы сравнили уровни miR-3648 в семи клеточных линиях GC и нормальной клеточной линии GES-1 желудка. Результат показал, что уровни miR-3648 были ниже во всех клетках GC по сравнению с клетками GES-1 ( P < 0,001, рис. 1E). Это согласуется с нашими клиническими данными о том, что ткани желудка человека имеют низкие уровни миР-3648 (рис. 1C, D). В частности, уровни экспрессии миР-3648 значительно снизились в образцах ткани GC от пациентов с распространенным раком (стадия III + IV) по сравнению с образцами ткани GC от пациентов с ранней стадией рака (стадия I + II) (рис. 1F). В-четвертых, экспрессия miR-3648 была ниже при более глубокой инвазии GC (T3-4 по сравнению с T1-2), что позволяет предположить, что ее дефицит может способствовать инвазивности клеток GC (Fig. 1G). В-пятых, мы выполнили ISH желудочных образцов. Результаты также показали, что экспрессия миР-3648 в GC была значительно ниже, чем в нормальной слизистой оболочке желудка (рис. 1H и дополнительная рис. 2)
В совокупности эти данные свидетельствуют о том, что экспрессия миР-3648 подавляется в тканях РЖ и может быть задействована в качестве микроРНК-супрессора опухолей при РЖ.
Экзогенная миР-3648 подавляет злокачественное биологическое поведение клеток РЖ in vitro.
образцы тканей. Мы показали, что низкая экспрессия miR-3648 коррелирует с дифференцировкой ( P < 0,05), глубина инвазии ( P < 0,001), метастазы в лимфатические узлы (присутствуют или отсутствуют, P = 0,001) и стадия TNM (I–II против III–IV, P < 0,001), но не коррелировал с возрастом, полом и размером опухоли пациентов ( P > 0,05). Эти исследования показывают, что низкий уровень экспрессии миР-3648 связан со злокачественным прогрессированием у пациентов с РЖ (дополнительная таблица 4). Чтобы определить роль экспрессии miR-3648 в росте клеток и метастазировании в клетках GC, мы трансфицировали миметики miR-3648 или m-NC в клетки AGS и MKN45 или ингибитор miR-3648 или i-NC в клетки GES-1, подтверждение экспрессии миР-3648 с помощью количественной ОТ-ПЦР (рис. 2А). Затем мы исследовали, участвует ли миР-3648 в пролиферации клеток GC. Анализ проточной цитометрии показал, что сверхэкспрессия миР-3648 приводила к увеличению количества клеток в фазе G0/G1 клеточного цикла (фиг. 2В). Более того, экспрессия белков, связанных с клеточным циклом, таких как циклин D1, CDK4 и CDK6, значительно снижалась при обработке миметиками miR-3648, тогда как уровни циклина B1 оставались неизменными по сравнению с клетками AGS и MKN45, обработанными m-NC (рис. 2С). Кроме того, анализы CCK-8 показали, что обработка миметиками miR-3648 приводила к значительно более низким значениям оптической плотности при 450 нм, тогда как трансфекция клеток GES-1 ингибитором miR-3648 приводила к более высоким значениям OD, чем соответствующая группа NC ( Рис. 2D1, D2). Анализы образования колоний показали гораздо более низкое образование колоний в группе миметиков miR-3648, чем в группе m-NC. Напротив, трансфекция клеток GES-1 ингибитором miR-3648 имела противоположный эффект (рис. 2E1, E2). Кроме того, анализы EdU показали аналогичные результаты (рис. 2F1, F2).
A Клетки GC, трансфицированные миметиками miR-3648 или ингибиторами miR-3648 через 48 ч, и экспрессию miR-3648 оценивали с помощью qRT-PCR. *** P < 0,01; **** P < 0,001. Столбики погрешностей представляют собой среднее ± SD из трех независимых экспериментов. Значения P оценивали с помощью двустороннего непарного t -критерия Стьюдента. B Фазы клеточного цикла обработанных клеток оценивали с помощью проточной цитометрии. Представлен процент каждой фазы. ** P < 0,05; **** P < 0,001, двусторонний непарный критерий Стьюдента t -критерий. C Были приготовлены цельноклеточные лизаты клеток GC, и экспрессия белка была обнаружена вестерн-блоттингом. Рядом с полосами были отмечены значения шкалы серого. D – F Влияние миметиков или ингибиторов miR-3648 на пролиферацию клеток оценивали с помощью CCK-8 ( D ), образования колоний ( E ) и EdU (5-этинил-2´-дезоксиуридин). ( F ) анализ. *** P < 0,01 и **** P < 0,001; m-NC против миметиков и i-NC против ингибитора. G1 , G2 Анализы на заживление ран использовали для определения подвижности клеток GC после трансфекции миметиками или ингибиторами. Процент закрытия ран показан справа или под панелью. *** P < 0,01 и **** P < 0,001, м-НК против миметиков; ** P < 0,05 и **** P < 0,001, i-NC по сравнению с ингибитором. h2 , h3 Анализы инвазии проводили с использованием клеток, трансфицированных миР-3648. Количественные результаты показаны под панелью. **** P < 0,001, м-НК против миметиков; **** P < 0,001, i-NC по сравнению с ингибитором. Масштабные линейки, 50 мкм в F и H . Столбики погрешностей представляют собой среднее ± SD. Все эксперименты были повторены не менее трех раз с идентичными результатами. Значения P были оценены с использованием двустороннего непарного критерия Стьюдента т -тест.
Изображение с полным размером
Влияние миР-3648 на подвижность клеток и инвазию тестировали с использованием анализов заживления царапин и инвазии. Результаты показали, что эктопическая экспрессия миР-3648 значительно ингибировала способность к миграции и инвазии клеток AGS и MGN45, тогда как обработка ингибитором миР-3648 увеличивала способность к миграции и инвазии клеток GES-1 (рис. 2G1, G2, h2, h3). ).
В совокупности эти результаты in vitro позволяют предположить, что миР-3648 подавляет рост, миграцию и инвазию клеток GC.
miR-3648 непосредственно нацелена на 3′-UTR FRAT1 и FRAT2
Для определения молекулярных механизмов, с помощью которых miR-3648 регулирует рост и метастазирование клеток GC, были использованы два общедоступных биоинформационных алгоритма (mirDIP и DIANA) и микроРНК на основе секвенирования РНК. -3648 сигнатур использовали для анализа генов-мишеней миР-3648. Была взята коллекция генов с абсолютным кратным изменением> 1,2 в клетках AGS и сопоставлена с результатами двух опубликованных механизмов прогнозирования целей, mirDIP и DIANA. Результаты показали, что 102 гена были подавлены в клетках со сверхэкспрессией miR-3648 по сравнению с клетками m-NC (рис. 3A, B). Анализ обогащения этих 102 генов был проведен в Metascape (https://metascape.org/). Наиболее значимым биологическим процессом GO был сигнальный путь Wnt, который был обогащен в общей сложности 12 генами, включая CCND1, CELSR2, MST1R, SKI, TLE2, FRAT1, FRAT2, KREMEN1, ZBED3, LZTS2, NRARP и NLGN2 (дополнительная рис. 3A). ). Среди этих кандидатов CCND1 [27], MST1R [28], SKI [29], FRAT1 и FRAT2 [30] действуют как онкогены при РЖ. Таким образом, мы провели количественную ПЦР для обнаружения экспрессии мРНК CCND1, MST1R, SKI, FRAT1 и FRAT2 после увеличения или ингибирования экспрессии miR-3648. Как показано на дополнительных рисунках 3B, C, FRAT1 и FRAT2 имеют наиболее очевидные изменения. Таким образом, FRAT1 и FRAT2 были идентифицированы как потенциальные мишени.
A На трехсторонней диаграмме Венна указано количество генов, которые перекрываются в двух общедоступных алгоритмах биоинформатики (mirDIP и DIANA) и сигнатуре miR-3648 на основе секвенирования РНК. B Тепловая карта была основана на 102 генах-кандидатах, которые были подавлены в клетках AGS. Красный цвет представляет уровень экспрессии выше среднего, зеленый цвет представляет экспрессию ниже среднего. Выделено красной рамкой. Уровень экспрессии белка C miR-3648 и уровень экспрессии белка D FRAT1/FRAT2 в двенадцати свежесобранных тканях слизистой оболочки желудка с использованием анализа qRT-PCR и вестерн-блоттинга. Столбики погрешностей представляют собой среднее ± SD из трех независимых экспериментов. E Предполагаемая связывающая миР-3648 последовательность в пределах 3′-UTR мРНК FRAT1 или FRAT2. Указаны мутации в комплементарном сайте для исходной области миР-3648 в 3′-UTR гена FRAT1 или FRAT2. F Репортерные векторы люциферазы, содержащие 3′-UTR FRAT1 или FRAT2 дикого типа (wt) или мутантные (mut) 3′-UTR, были сконструированы и котрансфицированы миметиками miR-3648 в клетки AGS и MKN45. Репортерные анализы люциферазы использовали для определения того, связывается ли miR-3648 напрямую с 3′-UTR FRAT1 или FRAT2. * P > 0,05, *** P < 0,01 и **** P < 0,001, м-НК против миметиков. Столбики погрешностей представляют собой среднее ± SD из трех независимых экспериментов. Значения P были оценены с использованием двустороннего непарного критерия Стьюдента т -тест. G Вестерн-блот анализ уровней FRAT1 или FRAT2 в эпителиальных клетках желудка, обработанных миметиком или ингибитором миР-3648.
Изображение в полный размер
Чтобы определить корреляцию между миР-3648 и экспрессией потенциальных генов-мишеней FRAT1 или FRAT2, мы исследовали экспрессию миР-3648 и FRAT1 или FRAT2 в 12 парах тканей GC человека и сопоставили нормальную слизистую оболочку желудка с помощью микроРНК. и вестерн-блот анализы. Как показано на рис. 3C, D, у большинства пациентов наблюдались более низкие уровни miR-3648, но более высокие уровни белков FRAT1 и FRAT2 в раковых тканях (T) по сравнению с нормальной слизистой оболочкой желудка (N). Корреляционный анализ Спирмена (дополнительный рисунок 3D) выявил отрицательную корреляцию между miR-3648 и FRAT1 (дополнительный рисунок 3D1) и между miR-3648 и FRAT2 (дополнительный рисунок 3D2), а также положительную корреляцию между FRAT1 и FRAT2 (дополнительный рисунок). 3Е) выражение.
Мы провели анализ репортера люциферазы, чтобы определить, действительно ли FRAT1 или FRAT2 регулируется miR-3648. Для этого мы сконструировали 3′-UTR репортерные плазмиды, кодирующие полноразмерные сайты связывания miR-3648 дикого типа (wt) или мутантные (mut) для FRAT1 или FRAT2 (рис. 3E). Мы обнаружили, что miR-3648 репрессирует репортерную активность плазмиды wt-FRAT1-3′-UTR или wt-FRAT2-3′-UTR, но не мут-FRAT1-3′-UTR или mut-FRAT2-3′. -UTR плазмида (фиг. 3F).
Затем вестерн-блот-анализ показал, что сверхэкспрессия миР-3648 подавляла экспрессию эндогенных FRAT1 или FRAT2 в клетках AGS и MKN45, тогда как блокирование экспрессии миР-3648 приводило к усилению эндогенной экспрессии FRAT1 или FRAT2 в клетках GES-1 (рис. 3G).
В совокупности эти результаты позволяют предположить, что miR-3648 может подавлять экспрессию FRAT1 и FRAT2 путем непосредственного нацеливания на ее 3′-UTR.
FRAT1 и FRAT2 участвуют в опосредованном miR-3648 ингибировании инвазии и метастазирования клеток GC
FRAT1 и FRAT2 являются важными эффекторами некоторых микроРНК при раке человека [31,32,33]; следовательно, необходимо дальнейшее изучение биологических эффектов miR-3648 на FRAT1 и FRAT2. Мы трансфицировали клетки AGS и MKN45 миметиками m-NC, miR-3648, вектором, плазмидой FRAT1 или плазмидой FRAT2 и котрансфицировали их miR-3648 + FRAT1 или miR-3648 + FRAT2. Анализы клеточных функций показали, что активация miR-3648 приводила к снижению способности к пролиферации, миграции и инвазии, тогда как эктопическая активация FRAT1 или FRAT2 плюс miR-3648 частично обращала влияние miR-3648 на рост, миграцию и инвазию клеток GC по сравнению с с контрольными клетками (дополнительная рис. 4A, B и рис. 4A, B).
A Способность к миграции и B способность к инвазии клеток, трансфицированных миметиками miR-3648 и/или экспрессионной плазмидой FRAT1 или FRAT2, определяли с использованием анализа заживления ран или анализа Transwell. ** P < 0,05; *** P < 0,01; **** P < 0,001. Результат представлен как среднее ± SD. Значения P были оценены с использованием двух — хвостатая непарная Стьюдента т -тест. Все эксперименты были повторены не менее трех раз с идентичными результатами. C Клетки GC MKN45 ортотопически трансплантировали в легкие бестимусных мышей, и мышей случайным образом разделили на семь групп следующим образом: m-NC, miR-3648, Vector, FRAT1, FTAT2, miR-3648 + FRAT1 и miR-3648. + группа FRAT1 ( n = 3 в каждой группе, была показана одна из трех бестимусных мышей). Столбики погрешностей представляют собой среднее ± SD. Значения P были оценены с использованием двух — хвостатая непарная Стьюдента т -тест. D Мышей умерщвляли и метастатические раковые ткани окрашивали гематоксилином и эозином (H&E). E Были показаны репрезентативные изображения метастатических очагов в легких. Подсчитывали количество метастатических очагов в легких. ** P < 0,05; *** P < 0,01; **** P < 0,001. F Иммуногистохимическое (IHC) окрашивание экспрессии MMP2 и MMP9. Шкала баров, 50 мкм в B , 200 мкм в D и 100 мкм в F .
Изображение полного размера
Предыдущие отчеты показали, что FRAT1 или FRAT2 связаны с инвазией и подвижностью раковых клеток. Таким образом, мы определили, участвует ли FRAT1 или FRAT2 в опосредованном miR-3648 ингибировании метастазирования клеток GC in vivo. Клетки MKN45, трансфицированные Lv-m-NC, Lv-miR-3648, Lv-Vector, Lv-FRAT1, Lv-FRAT2, Lv-miR-3648 + FRAT1 или Lv-miR-3648 + FRAT2, инъецировали в хвост. вена голых мышей (рис. 4C и дополнительная рис. 4C). Гистологический анализ срезов легких, окрашенных H&E, подтвердил наличие метастазов в легких (рис. 4D). Мы показали, что количество метастатических локусов было явно уменьшено у мышей, которым инъецировали клетки, экспрессирующие miR-3648, по сравнению с теми, которым вводили клетки m-NC, тогда как клетки, экспрессирующие FRAT1 или FRAT2, образовывали больше метастатических узелков, чем FRAT1/miR-3648 или FRAT1. /miR-3648, экспрессирующие совместно и вектор-экспрессирующие клетки (фиг. 4E). ММП2 и ММП9представляют собой желатиназы семейства матриксных металлопротеиназ, которые изначально были связаны с инвазивными и метастатическими свойствами опухолевых клеток благодаря их способности разрушать основные белковые компоненты внеклеточного матрикса и базальных мембран [34]. Таким образом, иммуногистохимическое (IHC) окрашивание с использованием антител против MMP2 и против MMP9 дополнительно подтвердило метастатическую способность различных групп (рис. 4F). В совокупности наши данные демонстрируют, что миР-3648 подавляет инвазию и метастазирование GC путем подавления FRAT1 или FRAT2.
Взаимодействие FRAT1 и FRAT2 способствует инвазии и метастазированию в клетки РЖ
Гены FRAT1 и FRAT2, ассоциированные с раком, сгруппированы в хромосоме человека в локусе 10q24 [30]. Поэтому мы сначала исследовали, существует ли корреляция между экспрессией FRAT1 и FRAT2 в клетках GC. Мы исследовали взаимодействующие белки FRAT1 в базе данных GeneMANIA (http://genemania. org/), и результаты показали, что белок FRAT1 может связываться с белком FRAT2. Корреляционный анализ Пирсона показал положительную корреляцию между FRAT1 и FRAT2 с использованием базы данных GEPIA (http://gepia.cancer-pku.cn/) (дополнительная рис. 5A). Затем мы исследовали экспрессию FRAT1 и FRAT2 в 82 тканях GC человека и обнаружили положительную корреляцию между FRAT1 и FRAT2 с помощью количественной ПЦР (дополнительная рис. 5B).
Для дальнейшей проверки физического взаимодействия между FRAT1 и FRAT2 in vitro мы трансфицировали FRAT1, помеченный Flag, в клетки AGS и MKN45 и оценили, взаимодействует ли белок FRAT1 с FRAT2. Мы показали, что антитело против флага (FRAT1) может совместно иммунопреципитировать FRAT2 из клеточных экстрактов (рис. 5В1). Точно так же антитело против HA (FRAT2) может коиммунопреципитировать FRAT1 (фиг. 5B2). Конфокальная флуоресцентная микроскопия показала, что совместная локализация двух белков была очевидна на объединенных изображениях (рис. 5C).
Рис. 5: Синергизм FRAT1 с FRAT2 способствует инвазии и метастазированию опухоли. A Потенциальные партнеры по связыванию FRAT2 были предсказаны с использованием базы данных GeneMANIA. Красные прямоугольники представляют белок-белковые взаимодействия. B1 , B2 Взаимодействие между FRAT1 и FRAT2 в клетках GC. Меченую флагом FRAT1 ( B1 ) или HA-меченную FRAT2 ( B2 ) плазмиду трансфицировали в клетки. Иммунопреципитацию проводили с меченым Flag антителом FRAT1 или HA-меченым FRAT2, а иммуноглобулин G нормальной мыши до иммунизации (nm IgG) использовали в качестве контроля. Вестерн-блот-анализ проводили с использованием антител против FRAT2 (α-FRAT2) или против FRAT1 (α-FRAT1). C Двойное окрашивание FRAT1 (зеленый) и HA-FRAT2 (красный) в клетках GC визуализировали с помощью конфокальной микроскопии; ядра были контрастно окрашены Hoechst 33258. D Уровни белка FRAT2 в клетках AGS и MKN45 после четырех обработок [Scrambled (Scr) siRNA, FRAT2 siRNA 1, FRAT2 siRNA 2 и FRAT2 siRNA 3] определяли вестерн-блот-анализом. E In vitro инвазивную способность клеток AGS и MKN45 оценивали с помощью анализа Транвелла. миРНКп, миРНКпул. **** Р < 0,001. Столбики погрешностей представляют собой среднее ± SD. Значения P оценивали с помощью двустороннего непарного t -критерия Стьюдента. F Мышам ортотопически трансплантировали клетки MKN45 ( n = 3 в каждой группе, показана одна из трех голых мышей). Столбики погрешностей представляют собой среднее ± SD. Значения P оценивали с помощью двустороннего непарного t -критерия Стьюдента. G Метастатические раковые ткани окрашивали H&E. Ч Подсчитано количество метастатических очагов в легких. ** P < 0,05; *** P < 0,01. I IHC-окрашивание экспрессии MMP2 и MMP9. Шкала баров соответствует 20 мкм для C , 50 мкм для E , 200 мкм для G и 100 мкм для I .
Изображение полного размера
Мы синтезировали три пары siРНК FRAT2 в клетках GC и подтвердили их с помощью вестерн-блоттинга (рис. 5D). Чтобы показать, проявляет ли FRAT1 синергизм с FRAT2 во время метастазирования и инвазии, инвазию GC-клеток in vitro определяли с использованием анализов заживления царапин и трансвелл. Мы показали, что принудительная экспрессия FRAT1 увеличивала способность к миграции и инвазии по сравнению с обработкой одним вектором, тогда как подавление FRAT2 в клетках со сверхэкспрессией FRAT1 снижало клеточную подвижность и потенциал инвазии клеток со сверхэкспрессией FRAT (дополнительная рис. 5C и рис. 5Е).
Для дальнейшего подтверждения синергизма FRAT1 с FRAT2 мы также синтезировали три siРНК FRAT1. Эффекты этих трех siRNA FRAT1 были подтверждены вестерн-блоттингом (дополнительная фиг. 6A). Мы оценили способность клеток GC к миграции и инвазии, используя тесты на заживление ран и анализы инвазии через лунки. Мы обнаружили, что подавление экспрессии FRAT1 или FRAT2 соответственно может ингибировать способность к миграции и инвазии клеток GC, в то время как подавление экспрессии FRAT1 и FRAT2 одновременно значительно усиливает ингибирующий эффект (дополнительная фиг. 6B, C).
Чтобы устранить функциональные последствия взаимодействия FRAT1 с FRAT2 при метастазировании опухоли, мы инокулировали клетки MKN45, трансдуцированные Lv-vector, Lv-FRAT1, Lv-shRNA, Lv-FRAT2 shRNA2 или Lv-FRAT1 + FRAT2 shRNA2, в BALB/c. Мыши -nu/nu (рис. 5F и дополнительная рис. 6D). Наличие метастазов ГК в легкое было подтверждено гистологическим анализом (рис. 5G). В группе Lv-FRAT1 были обнаружены более крупные метастатические узлы в легких, чем в группе Lv-vector. Напротив, инокуляция клетками Lv-FRAT1-FRAT2-shRNA2 обращала вспять эффекты, наблюдаемые у мышей, инокулированных клетками Lv-FRAT1 (фиг. 5H). Анализы IHC показали, что сверхэкспрессия FRAT1 приводит к значительному усилению метастатических маркеров MMP2 и MMP9., тогда как потеря FRAT2 в клетках со сверхэкспрессией FRAT1 вызывала снижение экспрессии MMP2 и MMP9 (фиг. 5I).
В совокупности эти данные свидетельствуют о том, что ось FRAT1-FRAT2 способствует прогрессированию рака и метастазированию.
миР-3648 ингибирует метастазирование клеток GC путем инактивации пути передачи сигнала Wnt/β-катенин путем подавления FRAT1/FRAT2
Предыдущие исследования показали, что FRAT1 и FRAT2 являются важными участниками пути передачи сигнала Wnt/β-катенина [ 35,36,37,38]. Поэтому мы сначала временно котрансфицировали клетки GC вектором FRAT1 и/или siRNA FRAT2 и репортерными плазмидами pTOPFLASH или pFOPFLASH. Репортерный анализ TOPFLASH/FOPFLASH представляет собой метод относительного количественного обнаружения для измерения активности сигнального пути Wnt/β-катенина [39]., 40] и проводили через 48 ч после обработки в клетках GC. Мы обнаружили, что эктопическая экспрессия FRAT1 значительно усиливала активность β-катенин-зависимой люциферазы, тогда как подавление FRAT2 в клетках со сверхэкспрессией FRAT1 обращало усиленный эффект (Fig. 6A). Во-вторых, мы проверили, регулирует ли miR-3648 сигнальный путь Wnt/β-catenin. Результаты показали, что miR-3648 является негативным регулятором передачи сигналов Wnt/β-catenin (Fig. 6B1, B2).
A Клетки GC котрансфицировали вектором, или FRAT1, или Scr siRNA, или FRAT2 siRNAp, или FRAT1 + FRAT2 siRNAp, и репортерную плазмиду TOP/FOPflash измеряли через 48 ч и определяли активность люциферазы. Нормализованная активность люциферазы Renilla, используемая в качестве внутреннего контроля. *** P < 0,01; **** P < 0,001. Двусторонний непарный тест Стьюдента t -тест. Клетки GC B1 , B2 miR-3648 трансфицировали репортерной плазмидой TOP/FOPflash и через 48 ч определяли репортерную активность. **** P < 0,001; **** P < 0,001. Двусторонний непарный тест Стьюдента t -тест. C Клетки GC котрансфицировали m-NC, или miR-3648, или Vector, или FRAT1, или FRAT2, или miR-3648 + FRAT1, или miR-3648 + FRAT2 и репортерной плазмидой TOP/FOPflash. Затем активность люциферазы тестировали через 48 часов. *** P < 0,01; **** P < 0,001. Двусторонний непарный тест Стьюдента t -тест. D Клетки GC котрансфицировали и через 48 часов измеряли активность люциферазы. В качестве внутреннего контроля использовали нормализованную активность люциферазы Renilla. *** P < 0,01; **** P < 0,001. Двусторонний непарный тест Стьюдента t -тест. E Клетки GC котрансфицировали 100 нг репортера люциферазы TopFlash или FOPFlash и m-NC + ДМСО, или miR-3648 + ДМСО, или m-NC + KYA1797K, или miR-3648 + KYA1797K, затем обрабатывали в течение 48 ч. . Измеряли активность люциферазы. *** P < 0,01; **** P < 0,001. Двусторонний непарный тест Стьюдента t -тест. F , G Эффекты m-NC + ДМСО или miR-3648 + ДМСО, m-NC + KYA1797K или miR-3648 + KYA1797K на миграцию и инвазию клеток GC с использованием трансвеллов и анализа заживления ран. ** P < 0,05; *** P < 0,01; **** P < 0,001. Двусторонний непарный тест Стьюдента t -тест. Все эксперименты повторяли не менее трех раз. h2 , h3 Вестерн-блот анализ экспрессии десяти белков в клетках GC, трансфицированных миметиками miR-3648 или ингибитором miR-3648. В качестве контроля использовали клетки, трансфицированные контрольными (NC) плазмидами. Данные денситометрии, представленные под полосами, представляют собой кратное изменение по сравнению с контролем. Шкала баров, 50 мкм в Ф .
Изображение с полным размером
В-третьих, мы стремились проверить, опосредует ли FRAT1 или FRAT2 активацию сигнального пути Wnt/β-catenin в клетках GC с низкой экспрессией miR-3648. Принудительная экспрессия FRAT1 или FRAT2 приводила к повышению активности β-catenin-зависимой люциферазы, тогда как miR-3648 подавляла ее. Кроме того, совместная экспрессия FRAT1, FRAT2 и миР-3648 обращала вспять эффект сверхэкспрессии миР-3648 (фиг. 6С). Более того, повышенная активация сигнального пути Wnt/β-катенина в клетках AGS и MKN45, трансфицированных ингибитором miR-3648, в значительной степени устранялась нокдауном FRAT1 или FRAT2 (Fig. 6D).
KYA1797K, небольшая молекула, ингибирует сигнальный путь Wnt/β-catenin [41, 42]. Затем мы определили, подавляет ли KYA1797K β-катенин-зависимую транскрипционную активность в клетках AGS и MKN45 посредством miR-3648. Мы показали, что миР-3648 или KYA1797K ослабляли активность люциферазы по сравнению с группой m-NC + ДМСО. Более того, группы miR-3648 и KYA1797K имели более низкие уровни транскрипционной активности TOPFLASH, чем группы m-NC + ДМСО или m-NC + KYA1797K (рис. 6E).
Для изучения эффекта миР-3648 или миР-3648 в сочетании с KYA1797K на миграцию и инвазию клеток, мы выполнили анализ трансвелла и заживление царапин в клетках GC соответственно. Результаты показали, что сверхэкспрессия miR-3648 или KYA1797K ингибировала миграцию и инвазию клеток GC по сравнению с клетками, обработанными m-NC + ДМСО. Кроме того, аддитивный эффект miR-3648 и KYA1797K снижал миграционную и инвазивную способность клеток РЖ по сравнению с обработанными группами m-NC + ДМСО или m-NC + KYA1797K (рис. 6F, G).
Белки FRAT1, FRAT2, GSK3b, β-катенин, TCF-4, MMP7, CD44, c-Myc, c-JUN и HIF-1a являются ключевыми участниками сигнального пути клеток Wnt/β-катенин, в котором отклонения в их экспрессии были связаны со злокачественной трансформацией клеток [12, 14, 18, 35, 36, 38, 43, 44]. Поэтому экспрессию этих десяти ключевых белков измеряли с помощью вестерн-блоттинга в клетках, обработанных m-NC и miR-3648 mimic или обработанных ингибитором miR-3648. Репрезентативные блоты для клеток GC показаны на рис. 6h2, h3. Относительную экспрессию этих белков далее рассчитывали путем нормализации по экспрессии GAPDH. Результаты показали, что эти десять белков показали значительно аномальные паттерны экспрессии в клетках, обработанных миметиком miR-3648 или ингибитором miR-3648, по сравнению с контрольными клетками (рис. 6h2, h3).
Эти результаты демонстрируют, что miR-3648 инактивирует сигнальный путь Wnt/β-catenin путем нацеливания на FRAT1 и FRAT2, влияя на миграцию и инвазию клеток GC.
miR-3648 напрямую регулируется фактором транскрипции c-Myc
Упомянутые выше результаты и предыдущие отчеты показали, что большое количество генов-мишеней, таких как c-Myc [13], MMP7 [45], циклин D1 [16, 17], c-JUN [15, 46] и HIF-1α [18] в сигнальном пути Wnt/β-catenin были идентифицированы. Эктопическая экспрессия miRNAs также регулируется генами фактора транскрипции, участвующими в сигнальном пути Wnt/β-catenin. Поэтому мы исследовали, могут ли факторы транскрипции c-Myc, c-JUN и HIF-1α регулировать уровни экспрессии miR-3648 в клетках GC посредством регуляторной петли обратной связи. Мы показали, что экспрессия miR-3648 оставалась неизменной при сверхэкспрессии c-Jun и HIF-1a в клетках GC. Напротив, эктопическая экспрессия c-Myc приводила к снижению экспрессии miR-3648 (фиг. 7А). Более того, сверхэкспрессия c-Myc приводила к снижению экспрессии первичного транскрипта miR-3648 (pri-miR-3648) в клетках GC (фиг. 7B). Затем мы сфокусировались непосредственно вверх по течению в области 1 т.п.н. сайта начала транскрипции миР-3648 и идентифицировали два предполагаемых сайта связывания c-Myc (рис. 7C) с по меньшей мере 84% идентичностью с консенсусными последовательностями c-Myc ( рис. 7D), которые располагались на отметке –189до –177 п.н. (сайт 1) и от –274 до –262 п.н. (сайт 2) в промоторе miR-3648.
A , B Сверхэкспрессия c-Myc, или HIF-1a, или JUN, или вектора трансфицировали в клетки AGS и MKN45. Относительные уровни экспрессии c-Myc, или HIF-1a, или JUN, или зрелой miR-3848 ( A ), или pri-miR-769 ( B ) были обнаружены в клеточных линиях AGS и MKN45 с помощью анализа qRT-PCR. . * Р > 0,05; *** P < 0,01; **** P < 0,001. Двусторонний непарный тест Стьюдента t -тест. C Мотив связывания транскрипционного фактора c-Myc был предсказан с использованием базы данных Jaspar. D Репортерные конструкции люциферазы (Luc) содержали промотор miR-3648 с двумя потенциальными сайтами связывания c-Myc. E Анализ ChIP-qPCR продемонстрировал прямое связывание c-Myc с промотором miR-3648 в клетках AGS и MKN45. Обогащение генами количественно определяли по сравнению с входным контролем с помощью количественной ПЦР с использованием праймеров, специфичных для промоторных областей миР-3648. Положительным контролем служили праймеры для ПЦР, расположенные в экзоне 3 RPL30, и антитело против гистона h4. Праймеры использовали для амплификации обслуживаемой области, содержащей в качестве фона удаленный вышестоящий промотор miR-3648. Результаты показаны как кратное изменение значения количественной ПЦР по сравнению с IgG. Двухвостая непарная Студенческая т -испытание; * P > 0,05 и **** P < 0,001. F c- Myc трансрепрессирует активность промотора miR-3648 в клетках AGS и MKN45. Конструкцию промотора miR-3648 котрансфицировали c-Myc или вектором и определяли относительную активность люциферазы. *** P < 0,01; **** P < 0,001. Двусторонний непарный тест Стьюдента t -тест. G , H Были проведены анализы миграции и инвазии клеток GC. *** P < 0,01; **** P < 0,001. Двусторонний непарный тест Стьюдента t -тест. Все эксперименты повторяли не менее трех раз. I Мышам ортотопически трансплантировали клетки MKN45 ( n = 3 в каждой группе, показана одна из трех голых мышей. *** P < 0,01; **** P < 0,001. непарный тест Стьюдента t -тест J Мышей умерщвляли и ткани метастатического рака окрашивали H&E K Подсчитано количество метастатических очагов в легких. *** P < 0,01; **** P < 0,001. L ИГХ-окрашивание экспрессии MMP2 и MMP9. Масштабные линейки, 50 мкм в H , 200 мкм в J и 100 мкм в L .
Изображение с полным размером
Чтобы определить, связывается ли c-Myc с промоторной областью miR-3648 in vivo, мы провели анализ ChIP в клетках GC. Расстояние между сайтами связывания c-Myc 1 и 2 составляло 73 п.н. Праймер для КПЦР нельзя сконструировать с помощью онлайн-программного обеспечения (праймер3 и праймер3плюс), потому что содержание GC достигает 84,9.% и трудно разделить сайты рядом друг с другом. Таким образом, мы объединили сайты связывания c-Myc 1 и 2 для разработки праймеров для ChIP-qPCR. Результаты показали, что промоторная область miR-3648, содержащая сайты 1 и 2, демонстрировала значительное обогащение после иммунопреципитации антителом против c-Myc (фиг. 7E). Затем мы клонировали промоторные области сайта c-Myc 1 или сайта 2 miR-3648 выше по течению в репортерную плазмиду люциферазы. Клетки AGS и MKN45 временно котрансфицировали вектором экспрессии c-Myc и репортерной плазмидой. Двойной люциферазный анализ показал, что активность сайтов 1 и 2 miR-3648 в клетках со сверхэкспрессией c-Myc снизилась примерно в 2 раза по сравнению с таковой в клетках, обработанных вектором (фиг. 7F). Эти результаты предполагают, что c-Myc трансрепрессирует промотор miR-3648 в клетках GC.
Чтобы определить, влияет ли miR-3648 на прогрессирование рака путем регуляции экспрессии c-Myc в клетках GC, мы трансфицировали c-Myc или вектор, miR-3648 или m-NC и c-Myc + miR-3648 в клетки AGS и MKN45. . По сравнению с контрольной группой сверхэкспрессия c-Myc повышала миграционную способность и инвазивность клеток GC in vitro, тогда как повторное введение миР-3648 не полностью обращало вспять эффекты c-Myc, стимулирующие миграцию и инвазивность (рис. 7G, H). .
Чтобы определить влияние miR-3648 на c-Myc-опосредованное метастазирование in vivo, клетки MKN45/Vector + m-NC, MKN45/c-Myc + m-NC и MKN45/c-Myc + miR-3648 были вводили в хвостовую вену голых мышей для выявления метастазов в легких (рис. 7I и дополнительная рис. 7). Наличие метастазов GC в легких было подтверждено окрашиванием гематоксилин-эозином (рис. 7J). Результаты показали, что количество метастатических узелков в легких было значительно увеличено у мышей, которым вводили клетки c-Myc, по сравнению с теми, которым вводили контрольные клетки. Однако в группе c-Myc/miR-3648 наблюдалось меньше метастатических узелков в легких, чем в группе c-Myc (рис. 7K). Экспрессия маркеров клеточного метастазирования MMP2 и MMP7 была подтверждена с помощью ИГХ (фиг. 7L).
Эти результаты позволяют предположить, что c-Myc снижает уровень экспрессии miR-3648 в клетках GC.
Уровни экспрессии miR-3648 отрицательно коррелируют с экспрессией c-Myc, FRAT1 и FRAT2 в образцах желудка человека
Чтобы продемонстрировать связь между уровнями экспрессии c-Myc, miR-3648, FRAT1 и FRAT2 в тканях GC, обнаружили экспрессию c-Myc, miR-3648, FRAT1 и FRAT2 в 82 человеческих тканях GC и парных нормальных тканях с помощью количественной ПЦР. Корреляционный анализ Спирмена показал отрицательную корреляцию между miR-3648 и FRAT1 (рис. 8A), между miR-3648 и FRAT2 (рис. 8B) и между c-Myc и miR-3648 (рис. 8C), а также положительную корреляцию между c-Myc и FRAT1 (фиг. 8D) и между c-Myc и FRAT2 (фиг. 8E).
A – E Корреляционный анализ Спирмена был использован для определения взаимосвязи между miR-3648 и FRAT1, между miR-3648 и FRAT2, между c-Myc и miR-3648, между miR-3648 и FRAT1 , а также между экспрессией c-Myc и FRAT2 в тканях 82 GC с использованием анализа qPCR. Уровни экспрессии F miR-3648, FRAT1, FRAT1 и c-Myc определяли с помощью анализов ISH и IHC. Эти цифры были представителями тканей желудка от двенадцати раковых и незлокачественных пациентов. Масштабные линейки, 50 мкм. G Гипотетическая модель, изображающая роли miR-3648, FRAT1, FRAT2 и c-Myc в клетках GC.
Изображение полного размера
Чтобы проверить, участвует ли вся сигнальная ось miR-3648/FRAT1-FRAT2/c-Myc в GC, мы оценили связь между уровнями экспрессии молекул, участвующих в этих путях, и их ассоциацией с Клинико-патологические особенности больных РЖ. Мы доказали, что существует корреляция между клинико-патологическими параметрами GC и экспрессией miR-3648 (дополнительная таблица 4). Мы обнаружили, что уровни экспрессии FRAT1, FRAT2 и c-Myc (дополнительные таблицы 5–7) в значительной степени коррелировали с метастазами GC в лимфатические узлы и стадией TNM.
Кроме того, мы обнаружили экспрессию четырех молекул с помощью анализов IHC и ISH. Уровни экспрессии FRAT1, FRAT2 и c-Myc были значительно выше, тогда как уровень экспрессии miR-3648 был значительно ниже в тканях GC, чем в парных нормальных тканях 12 образцов слизистой оболочки желудка (фиг. 8F).
Эти данные показывают, что экспрессия FRAT1, FRAT1 или c-Myc отрицательно коррелирует с уровнями экспрессии miR-3648 у пациентов с РЖ.
Обсуждение
миРНК играют решающую роль в канцерогенезе и могут регулировать множественные гены-мишени для модуляции метастатического прогрессирования при различных типах рака [3, 4, 6, 15, 19]. Однако биологические функции и механизмы, лежащие в основе действия miR-3648 при развитии и инвазии GC, еще не описаны. В этом исследовании мы определили функцию миР-3648 в подавлении роста и метастазирования опухоли GC. miR-3648 нацеливается на FRAT1 и FRAT2, которые, как известно, играют важную роль в сигнальном пути Wnt при прогрессировании GC. Более того, мы обнаружили, что промотор miR-3648 отрицательно регулируется геном-мишенью пути Wnt c-Myc. Таким образом, эти результаты предполагают, что петля отрицательной обратной связи miR-3648/FRAT1-FRAT2/c-Myc ингибирует инвазию и миграцию клеток GC.
В настоящее время хорошо известно, что микроРНК коррелируют с пролиферацией и агрессивностью опухоли при раке желудочно-кишечного тракта. Например, miR-500a-5p непосредственно нацеливается на HDAC2, который опосредует ингибирование пролиферации клеток CRC [47]. В этом исследовании мы впервые подтвердили, что миР-3648 значительно снижена при РЖ и что ее экспрессия связана с лучшим прогнозом у пациентов с РЖ. Сверхэкспрессия миР-3648 вызывает остановку клеточного цикла и ингибирует пролиферацию клеток GC. Более того, эктопическая экспрессия miR-3648 подавляла инвазию и миграцию клеток GC, что согласуется с предыдущими сообщениями о том, что miR-188-5p ингибирует ингибирование роста, подвижности и инвазии клеток GC [48]. В частности, ингибирование miR-3648 способствует развитию и прогрессированию GC, что позволяет предположить, что miR-3648 можно использовать в качестве биомаркера рака.
Гены FRAT1 и FRAT2 сгруппированы в хромосомном локусе человека 10q24.1, и оба гена ранее были идентифицированы как протоонкогены в различных опухолях [11, 30, 36], включая базальноподобный рак молочной железы [49]. ], рак яичников [37] и РЖ [30]. Однако остается неизвестным, участвует ли miR-3648 в регуляции экспрессии FRAT1 или FRAT2 при РЖ. В текущем исследовании мы идентифицировали FRAT1 и FRAT2 как основные нижележащие эффекторы miR-3648 в результате комбинированного анализа с использованием miRNA-seq и компьютерного предсказания мишени. Мы наблюдали, что миР-3648 негативно регулирует уровни белка FRAT1 или FRAT2 путем нацеливания на специфический сайт связывания в последовательности FRAT1 или FRAT2 3′-UTR. Между тем, сверхэкспрессия miR-3648 в клетках GC со сверхэкспрессией FRAT1 или FRAT2 регулировала клеточную пролиферацию и метастазирование как in vitro, так и in vivo. Таким образом, наше исследование предполагает, что миР-3648 является негативным регулятором клеточной пролиферации и метастазирования путем нацеливания на FRAT1 и FRAT2 при РЖ.
Ген FRAT1 кодирует белок из 279 — аминокислот, тогда как ген FRAT2 кодирует белок из 233 аминокислот, который на 77,3% идентичен FRAT1 [38]. Однако корреляция между экспрессией FRAT1 и FRAT2 при РЖ не выяснена. Здесь мы обнаружили, что FRAT1 физически связывается с FRAT2, чтобы способствовать метастазированию в клетках GC. Во-первых, мы продемонстрировали, что FRAT1 и FRAT2 могут совместно осаждаться в соответствии с базой данных GeneMANIA. Во-вторых, мы продемонстрировали положительную корреляцию между экспрессией FRAT1 и FRAT2 с использованием базы данных GEPIA и в 82 образцах тканей GC в нашей больнице. В-третьих, эксперименты по совместной иммунопреципитации и совместной локализации подтвердили, что FRAT1 взаимодействует с FRAT2, что согласуется с предыдущим отчетом van der Wal et al. что TMEM98 связывается с FRAT2 [50]. В-четвертых, FRAT1 взаимодействует с FRAT2, способствуя метастазированию опухоли in vitro и in vivo. Таким образом, мы предположили, что ось FRAT1-FRAT2 может играть роль в прогрессировании GC.
Передача сигналов Wnt представляет собой высококонсервативный сигнальный путь, который играет критическую роль в контроле эмбрионального развития и развития органов, а также прогрессии рака. Например, FZD7 является позитивным регулятором сигнального пути Wnt/β-catenin [51]. Его экспрессия была связана с плохим прогнозом у больных РЖ. Axin является негативным регулятором сигнального каскада Wnt. Его экспрессия подавляется в тканях РЖ и связана с метастазированием РЖ [52]. Однако роль FRAT1 и FRAT2 в модулировании сигнального пути Wnt в клетках GC полностью не раскрыта. В этом исследовании мы показали, что эктопическая экспрессия FRAT1 значительно повышает активность β-катенин-зависимой люциферазы, тогда как подавление FRAT2 в клетках со сверхэкспрессией FRAT1 обращает усиленный эффект, что согласуется с тем, что TCTP и Wnt-5a повышают β-катенин/TCF- 4 транскрипционной активности с помощью анализа гена отчета TOPflash/FOPflash [53, 54]. Следовательно, FRAT1 или FRAT2 по отдельности и FRAT1 синергизирует с FRAT2 для активации передачи сигналов Wnt/β-catenin в клетках GC.
Впоследствии мы исследовали, может ли miR-3648 ингибировать метастазирование опухоли GC путем модулирования сигнального пути FRAT1-FRAT2/Wnt/β-катенин в GC. Результаты анализов репортеров TOPFLASH/FOPFLASH показали, что FRAT1 или FRAT2 повышали активность Wnt/β-катенина, тогда как miR-3648 снижала активность Wnt/β-катенина в клетках GC. Более того, совместная трансфекция FRAT1 или FRAT2 и miR-3648 частично реверсировала эффект супрессии miR-3648 на активность Wnt/β-катенина. Вестерн-блоттинг показал, что миР-3648 подавляет Wnt 9.3697/ Экспрессия целевого гена β-катенина. Т.о., miR-3648 инактивирует сигнальный путь Wnt/β-catenin путем нацеливания на FRAT1 и FRAT2.
Недавние сообщения показали, что miRNAs в комбинации с антагонистами передачи сигналов Wnt/β-catenin отрицательно регулируют путь передачи сигналов Wnt/β-catenin при метастазировании опухоли [19, 55]. Соответственно, мы продемонстрировали, что клетки, обработанные миР-3648 и/или Wnt/β-катенин-ингибитором KYA1797K, демонстрировали более низкие уровни активности люциферазы, чем контроль. Кроме того, миР-3648 проявляет синергизм с KYA179.7K и ингибирует миграцию клеток GC и способность к инвазии по сравнению с контрольной группой. Таким образом, и миР-3648, и KYA1797K являются потенциальными терапевтическими стратегиями для пациентов с РЖ.
Все больше данных показывает, что нарушение регуляции экспрессии протоонкогена c-Myc посредством аберрантной передачи сигналов Wnt/β-catenin приводит к миграции и инвазии опухолевых клеток [13, 43, 56]. Как хорошо изученный фактор транскрипции, c-Myc может регулировать транскрипцию нескольких микроРНК путем связывания с их промоторными областями, тем самым модулируя прогрессирование рака. Например, c-Myc может ингибировать экспрессию miR-200b за счет связывания с промоторной областью гена miR-200b [57]. Ось c-Myc-miR-200b-PRDX2 регулирует прогрессирование CRC [58]. Здесь миР-3648 является прямой мишенью транскрипции c-Myc, о чем свидетельствует прямое связывание c-Myc с промотором miR-3648 и его трансрепрессия. Этот результат обеспечивает механизм, с помощью которого c-Myc ингибирует экспрессию miR-3648, регулируя метастазирование и инвазию GC. Кроме того, вышеупомянутые результаты показали, что miR-3648 нацелена на FRAT1 и FRAT2, и что FRAT1/FRAT2 положительно регулирует сигнальный путь Wnt и подавляет экспрессию c-Myc. Эти данные свидетельствуют о том, что miR-3648/FRAT1-FRAT2/c-Myc образует петлю отрицательной обратной связи, которая может ингибировать инвазию и метастазирование клеток GC.
В заключение, как показано в нашей рабочей модели на рис. 8G, мы показали, что miR-3648 инактивирует передачу сигналов Wnt/β-catenin путем прямого нацеливания на их регуляторы, FRAT1 и FRAT2. В свою очередь, инактивация передачи сигналов Wnt/β-catenin приводит к усилению экспрессии miR-3648 за счет уменьшения связывания c-Myc с промоторной областью miR-3648. Следовательно, miR-3648/FRAT1-FRAT2/c-Myc образует петлю отрицательной обратной связи, которая регулирует инвазию и метастазирование клеток GC. Наши результаты показали, что миР-3648 может быть новым прогностическим маркером и целью лечения РЖ.
Материалы и методы
Доступно в дополнительной информации.
Доступность данных
Наборы данных, использованные и проанализированные в ходе текущего исследования, можно получить у соответствующего автора по обоснованному запросу.
Ссылки
Li Q, Xu X, Su D, Zhou T, Wang G, Li Z. Долгосрочная выживаемость пожилого пациента с распространенным раком желудка после комбинированной терапии: отчет о клиническом случае и обзор литературы. БМК Рак. 2019;19:459.
ПабМед ПабМед Центральный Статья Google ученый
Ахади А. Нарушение регуляции миРНК как признак диагностики и прогноза рака желудка и их участие в механизме, лежащем в основе канцерогенеза и прогрессирования желудка.
Жизнь ИУБМБ. 2020; 72: 884–98.КАС пабмед Статья Google ученый
Ye G, Huang K, Yu J, Zhao L, Zhu X, Yang Q, et al. МикроРНК-647 нацелена на SRF-MYH9оси для подавления инвазии и метастазирования рака желудка. Тераностика. 2017;7:3338–53.
КАС пабмед ПабМед Центральный Статья Google ученый
Zhang P, Tang WM, Zhang H, Li YQ, Peng Y, Wang J и др. МиР-646 ингибировала пролиферацию клеток и индуцированное ЭМП метастазирование путем нацеливания на FOXK1 при раке желудка. Бр Дж Рак. 2017; 117: 525–34.
КАС пабмед ПабМед Центральный Статья Google ученый
Мин З., Сяомэн Л., Чжэн Л., Янге Д., Сюэцзяо Л., Лунвэй Л. и др. Асимметричная метилтрансфераза PRMT3 регулирует остеогенез мезенхимальных стволовых клеток человека посредством миР-3648.
Клеточная смерть Дис. 2019;10:581.ПабМед ПабМед Центральный Статья Google ученый
Xing R. миР-3648 способствует пролиферации клеток рака предстательной железы путем ингибирования аденоматозного полипоза толстой кишки 2. J Nanosci Nanotechnol. 2019;19:7526–31.
КАС пабмед Статья Google ученый
Sun W, Li S, Yu Y, Jin H, Xie Q, Hua X и др. Активация микроРНК-3648 приводит к подавлению TCF21, что приводит к инвазии и метастазированию рака мочевого пузыря человека. Молекулярные нуклеиновые кислоты. 2019;16:519–30.
КАС пабмед ПабМед Центральный Статья Google ученый
Karim R, Tse G, Putti T, Scolyer R, Lee S. Значение пути Wnt в патологии рака человека. Патология. 2004; 36: 120–8.
КАС пабмед Статья Google ученый
Katoh M, Katoh M. Транскрипционная регуляция WNT2B на основе баланса сигналов Hedgehog, Notch, BMP и WNT. Int J Oncol. 2009; 34:1411–5.
КАС пабмед Google ученый
van Amerongen R, Nawijn MC, Lambooij JP, Proost N, Jonkers J, Berns A. Онкопротеины Frat действуют на пересечении канонических и неканонических путей передачи сигналов Wnt. Онкоген. 2010;29:93–104.
ПабМед Статья Google ученый
Думпас Н., Лэмпарт Ф., Робинсон М.Д., Лентини А., Нестор С.Е., Канту С. и др. TCF/LEF-зависимая и независимая регуляция транскрипции генов-мишеней Wnt/бета-катенина. ЕМБО Дж. 2019;38:e98873.
He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT и др. Идентификация c-MYC как мишени пути APC. Наука. 1998; 281:1509–12.
КАС пабмед Статья Google ученый
Чжао М., Сюй П., Лю З., Жень И., Чен И., Лю И. и др. Двойная роль miR-374a с помощью модулированного c-Jun, соответственно, нацелена на CCND1-индуцирующий сигнал PI3K/AKT и PTEN-супрессирующий сигнальный сигнал Wnt/бета-катенин при немелкоклеточном раке легкого. Клеточная смерть Дис. 2018;9:78.
ПабМед ПабМед Центральный Статья Google ученый
Линь Дж., Дин С., Се С., Йи Р., Ву З., Луо Дж. и др. МикроРНК-4476 способствует прогрессированию глиомы через петлю положительной обратной связи miR-4476/APC/бета-катенин/c-Jun. Клеточная смерть Дис. 2020;11:269.
КАС пабмед ПабМед Центральный Статья Google ученый
Zhang W, Jiang B, Guo Z, Sardet C, Zou B, Lam CS, et al. Четыре с половиной белка LIM 2 способствуют инвазивному потенциалу и эпителиально-мезенхимальному переходу при раке толстой кишки. Канцерогенез. 2010;31:1220–9.
КАС пабмед Статья Google ученый
Сюй К., Чжан Ю., Юань З., Цю Ю., Ван Х., Фань Г. и др. Гипоксия индуцирует лекарственную устойчивость при колоректальном раке через петлю обратной связи HIF-1alpha/miR-338-5p/IL-6. Мол Тер. 2019;27:1810–24.
КАС пабмед ПабМед Центральный Статья Google ученый
Liu Q, Guan Y, Li Z, Wang Y, Liu Y, Cui R, et al. миР-504 подавляет мезенхимальный фенотип глиобластомы путем прямого нацеливания на FZD7-опосредованный путь Wnt-бета-катенина. J Exp Clin Cancer Res. 2019;38:358.
ПабМед ПабМед Центральный Статья Google ученый
Кодоло Г., Тоффолетто М., Чемелло Ф., Колетта С., Солер Тейксидор Г., Баттаджиа Г. и др. Helicobacter pylori ослабляет экспрессию HLA-II на макрофагах посредством повышающей регуляции микроРНК, нацеленных на CIITA. Фронт Иммунол. 2019;10:2923.
КАС пабмед Статья Google ученый
Lu J, Wang YH, Huang XY, Xie JW, Wang JB, Lin JX и др. circ-CEP85L подавляет пролиферацию и инвазию рака желудка, регулируя экспрессию NFKBIA посредством миР-942-5p. J Cell Physiol. 2020; 235: 6287–99.
КАС пабмед Статья Google ученый
Liu J, Wang X, Yang X, Liu Y, Shi Y, Ren J и др. миРНК423-5p регулирует пролиферацию и инвазию клеток путем нацеливания на фактор трилистника 1 в клетках рака желудка. Рак Летт. 2014;347:98–104.
КАС пабмед Статья Google ученый
Zhao C, Li Y, Chen G, Wang F, Shen Z, Zhou R. Сверхэкспрессия миР-15b-5p способствует метастазированию рака желудка путем регуляции PAQR3. Oncol Rep. 2017; 38:352–8.
КАС пабмед Статья Google ученый
Ибрагим Х.М., АбдЭльбари А.М., Мохамед С.Ю., Эльван А., Абдельхамид М.И., Ибрагим А. Прогностическое значение экспрессии циклина D1 и CD44 при аденокарциноме желудка. J Рак желудочно-кишечного тракта. 2019;50:370–9.
КАС пабмед Статья Google ученый
Zhou D, Zhu X, Wu X, Zheng J, Tou L, Zhou Y.
Эффект сплайсинга MST1R при раке желудка был усилен днРНК FENDRR. Эксперт Тер Мед. 2021;22:798.КАС пабмед ПабМед Центральный Статья Google ученый
Такахата М., Иноуэ Ю., Цуда Х., Имото И., Койнума Д., Хаяши М. и др. SKI и MEL1 совместно ингибируют трансформирующий бета-сигнал фактора роста в клетках рака желудка. Дж. Биол. Хим. 2009; 284:3334–44.
КАС пабмед Статья Google ученый
Saitoh T, Katoh M. FRAT1 и FRAT2, сгруппированные в области хромосомы 10q24.1 человека, активируются при раке желудка. Int J Oncol. 2001;19: 311–5.
КАС пабмед Google ученый
Werner TV, Hart M, Nickels R, Kim YJ, Menger MD, Bohle RM, et al. МиР-34a-3p изменяет пролиферацию и апоптоз клеток менингиомы in vitro и напрямую воздействует на SMAD4, FRAT1 и BCL2.
Старение. 2017; 9: 932–54.КАС пабмед ПабМед Центральный Статья Google ученый
Чжэн К., Чжоу С., Ю Дж., Ли К., Ван Х., Ли М. и др. Эпигенетическое молчание миР-490-3p способствует развитию агрессивного фенотипа колоректального рака посредством активации сигнального пути Wnt/бета-катенин. Рак Летт. 2016; 376: 178–87.
КАС пабмед Статья Google ученый
Jiang J, Yu C, Chen M, Zhang H, Tian S, Sun C. Снижение миР-29c усиливает миграцию клеток рака поджелудочной железы и фенотип, подобный стволовым клеткам. Онкотаргет. 2015;6:2767–78.
ПабМед Статья Google ученый
Folgueras AR, Pendas AM, Sanchez LM, Lopez-Otin C. Матриксные металлопротеиназы при раке: от новых функций к улучшенным стратегиям ингибирования. Int J Dev Biol.
2004;48:411–24.КАС пабмед Статья Google ученый
Сайто Т., Мориваки Дж., Койке Дж., Такаги А., Мива Т., Сиокава К. и др. Молекулярное клонирование и характеристика FRAT2, кодирующего позитивный регулятор сигнального пути WNT. Biochem Biophys Res Commun. 2001; 281:815–20.
КАС пабмед Статья Google ученый
Saitoh T, Mine T, Katoh M. Молекулярное клонирование и экспрессия протоонкогена FRAT1 при раке человека. Int J Oncol. 2002; 20: 785–9.
КАС пабмед Google ученый
Tian W, Lei N, Guo R, Yuan Z, Chang L. Длинная некодирующая РНК DANCR способствует росту рака шейки матки посредством активации сигнального пути Wnt/бета-катенина. Раковая ячейка Интерн. 2020;20:61.
ПабМед ПабМед Центральный Статья Google ученый
Ян С., Чжоу Ц., Ли М., Тонг С., Сунь Дж., Цин И. и др. Активация CYP2S1 оксалиплатином связана со статусом p53 в клеточных линиях колоректального рака. Научный доклад 2016; 6: 33078.
КАС пабмед ПабМед Центральный Статья Google ученый
Yang XZ, Cheng TT, He QJ, Lei ZY, Chi J, Tang Z и др. LINC01133 в виде цеРНК ингибирует прогрессирование рака желудка путем губчатой миР-106a-3p для регуляции экспрессии APC и пути Wnt/бета-катенин. Мол Рак. 2018;17:126.
ПабМед ПабМед Центральный Статья Google ученый
Ruan Z, Liang M, Lai M, Shang L, Deng X, Su X. KYA1797K подавляет PD-L1 в стволовых клетках рака толстой кишки, чтобы блокировать уклонение от иммунитета путем подавления сигнального пути бета-катенина/STT3. Int Immunopharmacol. 2020;78:106003.
КАС пабмед Статья Google ученый
Ван Б., Хсу С.Х., Ван Х., Кутай Х., Бид Х.К., Ю Дж. и др. Реципрокная регуляция микроРНК-122 и c-Myc при гепатоцеллюлярном раке: роль E2F1 и партнера по димеризации фактора транскрипции 2. Гепатология. 2014; 59: 555–66.
КАС пабмед Статья Google ученый
Fan H, Zhang M, Liu W. Гиперметилированный KCNQ1 действует как супрессор опухоли при гепатоцеллюлярной карциноме. Biochem Biophys Res Commun. 2018;503:3100–7.
КАС пабмед Статья Google ученый
Liu L, Ning Y, Yi J, Yuan J, Fang W, Lin Z и др. миР-6089/MYH9/бета-катенин/c-Jun петля отрицательной обратной связи ингибирует канцерогенез и прогрессирование рака яичников. Биомед Фармаколог. 2020;125:109865.
КАС пабмед Статья Google ученый
Пэн И, Шен С, Цзян Х, Чен З, У Дж, Чжу И и др. миР-188-5p подавляет пролиферацию и инвазию клеток рака желудка посредством нацеливания на ZFP91. Онкол рез. 2018;27:65–71.
ПабМед ПабМед Центральный Статья Google ученый
Zhou Y, Li C, Peng J, Luo P, Xie C, Liu S, et al. Регулятор сигнального пути WNT-FRAT2 влияет на онкогенез и прогноз базальноподобного рака молочной железы. Дж. Торак Дис. 2020;12:3478–87.
ПабМед ПабМед Центральный Статья Google ученый
ван дер Валь Т.
, Ламбуий Дж. П., ван Амеронген Р. TMEM98 представляет собой негативный регулятор опосредованной FRAT передачи сигналов Wnt/ss-катенина. ПЛОС Один. 2020;15:e0227435.ПабМед ПабМед Центральный Статья Google ученый
Kirikoshi H, Sekihara H, Katoh M. Активация Frizzled-7 (FZD7) при раке желудка человека. Int J Oncol. 2001;19:111–5.
КАС пабмед Google ученый
Маццони С.М., Фирон Э.Р. Варианты AXIN1 и AXIN2 при раке желудочно-кишечного тракта. Рак Летт. 2014; 355:1–8.
КАС пабмед ПабМед Центральный Статья Google ученый
Gu X, Yao L, Ma G, Cui L, Li Y, Liang W, et al. TCTP способствует пролиферации клеток глиомы in vitro и in vivo посредством усиленной транскрипции бета-катенина/TCF-4. Нейро Онкол. 2014;16:217–27.
КАС пабмед Статья Google ученый
Пакликова П., Радашкевич Т.В., Потесил Д., Харнос Дж., Здрахал З., Брыя В. Роль отдельных растрепанных паралогов человека в сигнальных путях Wnt. Сотовый сигнал. 2021;85:110058.
КАС пабмед Статья Google ученый
Ren L, Chen H, Song J, Chen X, Lin C, Zhang X и др. Опосредованное миР-454-3p подавление сигнальных антагонистов Wnt/бета-катенина способствует метастазированию рака молочной железы. Тераностика. 2019;9:449–65.
КАС пабмед ПабМед Центральный Статья Google ученый
Dang CV, Le A, Gao P. Энергетический метаболизм раковых клеток, индуцированный MYC, и терапевтические возможности. Клин Рак Рез. 2009 г.;15:6479–83.
КАС пабмед ПабМед Центральный Статья Google ученый
Lv Z, Wei J, You W, Wang R, Shang J, Xiong Y и др. Нарушение регуляторной петли c-Myc/miR-200b-3p/PRDX2 усиливает метастазирование опухоли и резистентность к химиотерапии при колоректальном раке. J Transl Med. 2017;15:257.
КАС пабмед ПабМед Центральный Статья Google ученый
Katoh M, Katoh M. Сигнальный путь WNT и сигнальная сеть стволовых клеток. Клин Рак Рез. 2007; 13:4042–5.
КАС пабмед Статья Google ученый
Штутман М., Журинский Дж., Симха И., Альбанезе К., Д’Амико М., Пестелл Р. и др. Ген циклина D1 является мишенью пути бета-катенин/LEF-1. Proc Natl Acad Sci USA. 1999;96:5522–7.
КАС пабмед ПабМед Центральный Статья Google ученый
Пэн Ю., Чжан С., Ма К., Ян Р., Цинь Ю., Чжао Ю. и др. МиРНК-194 активирует сигнальный путь Wnt/бета-катенин при раке желудка путем нацеливания на негативный регулятор Wnt, SUFU. Рак Летт. 2017; 385:117–27.
КАС пабмед Статья Google ученый
Ян З., Ши С., Ли С., Ван С., Хоу К., Ли З. и др. Длительная активация некодирующей РНК UCA1 способствует миграции устойчивых к гипоксии клеток рака желудка через ось miR-7-5p/EGFR. Разрешение ячейки опыта. 2018;368:194–201.
КАС пабмед Статья Google ученый
Zhang Y, Huang H, Zhang Y, Liao N. Комбинированное определение сывороточной экспрессии MiR-221-3p и MiR-122-5p в диагностике и прогнозировании рака желудка. J Рак желудка. 2019;19:315–28.
ПабМед ПабМед Центральный Статья Google ученый
Freemantle SJ, Portland HB, Ewings K, Dmitrovsky F, DiPetrillo K, Spinella MJ, et al. Характеристика и тканеспецифическая экспрессия человеческих GSK-3-связывающих белков FRAT1 и FRAT2. Ген. 2002; 291:17–27.
КАС пабмед Статья Google ученый
Yan G, Li S, Yue M, Li C, Kang Z. Лизиндеметилаза 5B подавляет CC хемокиновый лиганд 14, способствуя прогрессированию колоректального рака через путь Wnt/бета-катенин. Жизнь наук. 2021;264:118726.
КАС пабмед Статья Google ученый
Lv YF, Dai H, Yan GN, Meng G, Zhang X, Guo QN. Подавление кДНК 3 STF, подавляющей опухоль, способствует эпителиально-мезенхимальному переходу и метастазированию опухоли остеосаркомы посредством сигнального пути Wnt/GSK-3beta/beta-catenin/Snail. Рак Летт. 2016; 373:164–73.
КАС пабмед Статья Google ученый
Тан В., Чжоу В., Сян Л., Ву С., Чжан П., Ван Дж. и др. Сигнальная ось p300/YY1/miR-500a-5p/HDAC2 регулирует пролиферацию клеток при колоректальном раке человека. Нац коммун. 2019;10:663.
КАС пабмед ПабМед Центральный Статья Google ученый
Пан Ю, Лю Дж, Ли С, Сяо Г, Ван Х, Ян Г и др. Опосредованное MYC и DNMT3A метилирование ДНК репрессирует микроРНК-200b при тройном негативном раке молочной железы. J Cell Mol Med. 2018;22:6262–74.
КАС пабмед ПабМед Центральный Статья Google ученый
Скачать ссылки
Благодарности
Мы признательны за щедрую поддержку ключевой лаборатории гастроэнтерологии провинции Гуандун, отделение гастроэнтерологии, больница Наньфан, Южный медицинский университет. Это исследование было поддержано грантами Национального фонда естественных наук Китая (№ 81974448, 81772964, 82073066 и 82103152), Гуандунского фонда фундаментальных и прикладных фундаментальных исследований (№ 2020A1515110059). ), Гуандунский исследовательский центр желудочно-кишечных заболеваний (№ 2017B020209003), Гуандунский фонд медицинских исследований (№ B2019126), Шэньчжэньская комиссия по научным и технологическим инновациям (№ JCYJ20180306170328854), Шэньчжэньский фонд научно-технических инноваций (№ JCYJ20210324135005013) и Shenzhen Longang Фонд районной комиссии по инновациям в области науки и технологий (№ LGKCYLWS 2021000012).
Информация об авторе
Авторы и организации
Ключевая лаборатория гастроэнтерологии провинции Гуандун, отделение гастроэнтерологии, больница Наньфан, Южный медицинский университет, Гуанчжоу, 510515, Китай
Weimei Tang, Miaomiao Pei, Jiaying Li, Nanzhu Xu, Wushuang Xiao, Zhen Yu, Jieming Zhang, Linjie Hong, Weiyu Dai, Yizhi Xiao, Xiaosheng Wu, Guangnan Liu, Fachao Zhi, Jing Xiong, Aimin Li, Side Liu & Jide Wang
Отделение гематологии и онкологии, Международный онкологический центр, Больница общего профиля Шэньчжэньского университета, Xueyuan AVE 1098, Nanshan District, Shenzhen, Guangdong, 518000, P.
R. ChinaZheng Guo
Отделение гастроэнтерологии, Народная больница Longgang District, Shenzhen, 518172, China
Jianjiao Lin, Lide Xiao Lig, Ван
Отделение общей хирургии, больница Наньфан, Южный медицинский университет, Гуанчжоу, 510515, Китай
Гуоксин Ли
Центр клинической микроэкологии, больница Шэньчжэнь, Южный медицинский университет, Шэньчжэнь, 518000, Китай0910
Ye Chen
Department of Gastroenterology, Hexian Memorial Affiliated Hospital, Southern Medical University, Guangzhou, 511400, China
Hui Zhang
Authors
- Weimei Tang
View author publications
You can also search для этого автора в PubMed Google Scholar
- Miaomiao Pei
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Академия
- Jiaying Li
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Nanzhu Xu
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Wushuang Xiao
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Чжэнь Юй
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Jieming Zhang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Linjie Hong
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Zheng Guo
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Jianjiao Lin
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Weiyu Dai
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Yizhi Xiao
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Академия
- Xiaosheng Wu
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Guangnan Liu
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Fachao Zhi
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Guoxin Li
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Jing Xiong
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ye Chen
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Hui Zhang
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Li Xiang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Aimin Li
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Side Liu
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Академия
- Jide Wang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Взносы
Исследование было разработано и задумано JDW, SDL, AML, LX и HZ. Эксперименты in vitro проводились в основном с помощью WMT, MMP и JYL. NZX, WSX, ZY, JMZ и LJH проводили эксперименты in vivo. LX, ZG, JJL и WYD отвечали за анализ данных и статистический анализ. Образцы были собраны YZX, XSW и GNL. FCZ, GXL, JX и YC внесли свой вклад в техническую поддержку и редактирование рукописи.
Авторы переписки
Переписка с Ли Сян, Айминь Ли, Сиде Лю или Цзидэ Ван.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительная информация
Права и разрешения
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате, при условии, что вы укажете соответствующую ссылку на первоначальный автор(ы) и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения.